Capítulo 3
Metabolómica para aplicaciones comerciales
En este capítulo, aprenderás cómo la metabolómica puede aportar información funcional sobre los fenotipos de las enfermedades que los biomarcadores tradicionales no logran proporcionar, cómo puede identificar biomarcadores diagnósticos de gran eficacia que pasan desapercibidos para otras ciencias «ómicas», y cómo puede contribuir a la nutrición humana para prevenir enfermedades y mejorar la salud en general.
Visión general
La metabolómica se ha convertido en una herramienta de gran valor en diversos mercados aplicados, entre los que se incluyen el desarrollo de fármacos, la nutrición humana, la ganadería y el cuidado de mascotas. Los cambios metabólicos pueden ayudar a descubrir nuevas dianas terapéuticas y mecanismos de acción de los fármacos, identificar relaciones entre la dieta y los fenotipos de las enfermedades, y revelar biomarcadores que aportan información sobre la salud animal. En este artículo, analizamos algunos estudios que muestran cómo la metabolómica ha contribuido a resolver un problema o ha aportado conocimientos científicos clave sobre temas relacionados con determinados mercados aplicados.
Desarrollo de fármacos
Obtener información sobre la toxicidad inducida por fármacos que los biomarcadores tradicionales no han podido proporcionar
Antecedentes. La dislipidemia, es decir, un nivel anormalmente elevado de lipoproteínas de baja y muy baja densidad (LDL/VLDL) en la sangre, es un factor de riesgo importante de enfermedad cardiovascular y accidente cerebrovascular. Los agonistas del receptor de ácido nicotínico (NAR) tratan la dislipidemia activando los receptores de las células adiposas para inhibir el metabolismo de los triglicéridos y reducir los niveles circulantes de LDL y VLDL. En un estudio, los investigadores de Merck Research Laboratories estaban desarrollando un agonista del NAR (SCH 900424) para tratar la dislipidemia, pero en sus investigaciones identificaron una posible toxicidad inducida por el fármaco que deseaban comprender y controlar mejor [1].
Datos preliminares y objetivos del estudio. En los estudios preclínicos, el SCH 900424 provocó de forma inesperada una rápida morbilidad y una lesión renal aguda (LRA) en ratones, lo que sugiere que el fármaco afectaba tanto al sistema renal como al sistema nervioso central (SNC). Los marcadores tradicionales de la función renal, como la creatinina y el BUN, no eran lo suficientemente sensibles como para predecir o caracterizar la toxicidad inducida por el fármaco, y se desconocían los mecanismos subyacentes. Los objetivos de este estudio fueron identificar biomarcadores sensibles y tempranos de las toxicidades relacionadas con el SCH 900424 en ratones y dilucidar los mecanismos que provocaban la toxicidad.
Métodos. Se administraron a ratones CD-1 machos adultos dosis bajas, medias y altas de SCH 900424 o de un agonista de control. Se recogieron muestras de plasma, orina y cerebro 1, 4, 8 y 24 horas después de la administración. Las muestras se procesaron y se analizaron mediante metabolómica global para identificar alteraciones en las vías bioquímicas relevantes. A continuación, se utilizó la metabolómica dirigida para medir los compuestos bioquímicos de interés identificados en los análisis de descubrimiento global. Se analizaron tres matrices de muestras para garantizar un estudio mecánico exhaustivo de las patologías relacionadas tanto con la LRA como con el SNC.
Resultados. En plasma, el sulfato de 3-indoxilo (3IS), una toxina urémica de origen intestinal producida a partir del metabolismo del triptófano, se reveló como uno de los principales biomarcadores de la LRA inducida por SCH 900424. Este marcador aumentó significativamente en los ratones tratados en comparación con los controles y también se elevó antes y de forma más drástica que la creatinina o la urea (Figura 5, Dosis alta), lo que demuestra que el 3IS es un marcador más sensible de la IRA que los marcadores tradicionales de la función renal.
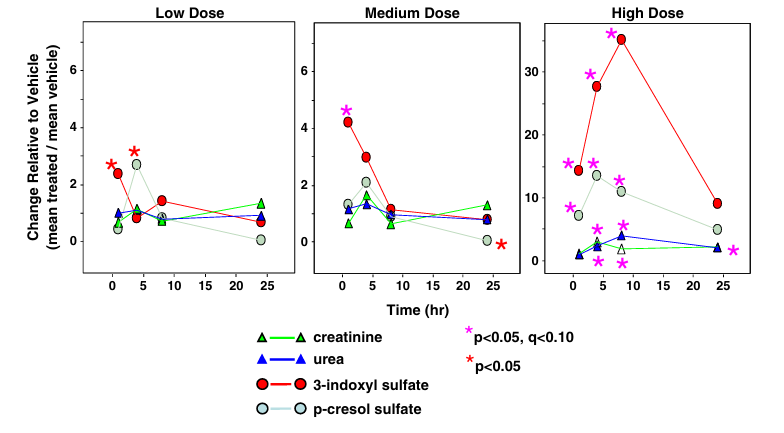
Figura 5. Variaciones porcentuales con respecto al vehículo en plasma para 3IS, PCS, urea y creatinina tras la administración de SCH 900424. Según el análisis metabolómico global exhaustivo, el 3IS y el sulfato de p-cresol se identificaron como los marcadores plasmáticos de toxicidad renal más estadísticamente significativos en este estudio y resultaron más sensibles que la urea y la creatinina. Imagen reproducida de Zgoda-Pols et al., Toxicol Appl Pharmacol, 2011, con licencia CC BY 4.0.
Los niveles plasmáticos de sulfato de p-cresol, una toxina urémica producida a partir del metabolismo de la tirosina y la fenilalanina, también se elevaron de forma significativa (Figura 5, dosis alta). Varios aminoácidos, entre ellos el triptófano, disminuyeron significativamente en plasma, mientras que muchos catabolitos del triptófano, como la kynurenina, el kynurenato y el indol-3-lactato, aumentaron de forma significativa (Figura 6). En la orina se observaron aumentos significativos de glucosa y aminoácidos. En condiciones normales, casi todos los aminoácidos y azúcares filtrados se reabsorben en los túbulos proximales. Sin embargo, una lesión renal alteraría la reabsorción y provocaría su excreción en la orina. La firma metabólica observada tanto en plasma como en orina apuntaba a una verdadera disfunción renal, más que a cambios metabólicos aislados.
Otro hallazgo importante del estudio fue el aumento significativo y dependiente de la dosis de 3IS en el tejido cerebral tras la administración de SCH 900424. Este aumento se produjo junto con una reducción significativa de los niveles de 3IS en la orina, lo que indica una alteración de la eliminación que conduce a su acumulación en el cerebro. En general, estos hallazgos sugieren que la acumulación de 3IS en el plasma y el cerebro podría contribuir no solo a la insuficiencia renal aguda, sino también a la toxicidad del SNC y a una rápida morbilidad en ratones.
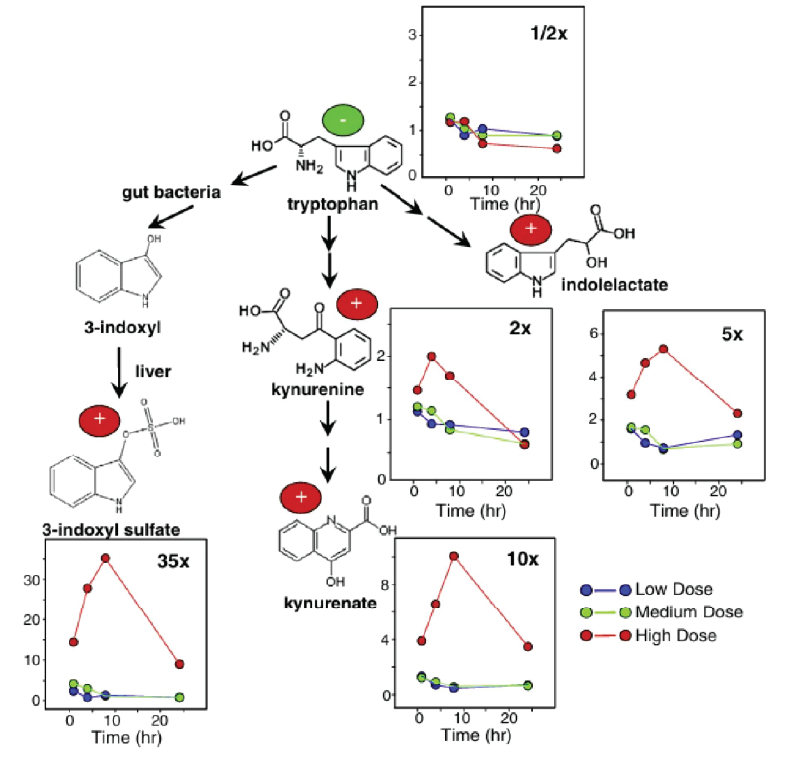
Figura 6. Vías catabólicas del triptófano con los cambios relativos respecto al vehículo para cada uno de los catabolitos. Mientras que los niveles de triptófano disminuyeron con el tiempo con las dosis más altas de SCH 900424, sus catabolitos, y especialmente el 3IS, aumentaron de forma espectacular. Imagen reproducida de Zgoda-Pols et al., Toxicol Appl Pharmacol, 2011, bajo licencia CC BY 4.0.
Conclusiones del estudio
- Los resultados de este estudio sugieren que el 3IS es un marcador precoz y más sensible de la toxicidad renal inducida por el SCH 900424 que los marcadores químicos tradicionales. En la práctica, esto podría mejorar la detección precoz de la nefrotoxicidad en el desarrollo de fármacos.
- La acumulación observada de toxinas urémicas de origen intestinal, la alteración del metabolismo del triptófano y los patrones de reducción de la excreción urinaria proporcionaron pruebas mecánicas de la acumulación de toxinas urémicas debida a la disfunción renal.
- La acumulación de 3IS en el tejido cerebral, y no solo en el plasma, sugiere una relación mecánica entre la insuficiencia renal y la rápida aparición de morbilidad en ratones; una hipótesis novedosa que podrá comprobarse en futuros estudios.
- Este estudio muestra cómo la metabolómica puede ayudar a convertir una observación fenotípica sin caracterizar en una historia de biomarcadores basada en los mecanismos subyacentes, lo que permite orientar la selección de candidatos a fármacos con mayor rapidez y precisión.
Cómo salvar un programa de desarrollo de fármacos
Antecedentes. La infección por C. difficile (ICD) provoca colitis inflamatoria y la consiguiente diarrea, lo que resulta muy debilitante para los pacientes. Incluso con tratamiento antibiótico, entre el 20 % y el 30 % de los pacientes sufren ICD recurrente debido a la alteración continuada de su microbioma. En un intestino sano, las bacterias intestinales inhiben el crecimiento y la germinación de C. difficile al convertir los ácidos biliares primarios conjugados en ácidos biliares secundarios y al producir ácidos grasos de cadena corta y media. Se ha demostrado que las esporas de la microbiota fecal producidas por miembros del filo Firmicutes recolonizan el intestino con bacterias que protegen contra la CDI recurrente. En un estudio, investigadores de Seres Therapeutics desarrollaron VOS (anteriormente SER-109), un producto bioterapéutico vivo basado en el microbioma, de administración oral, destinado al tratamiento y la prevención de la CDI recurrente [2].
Datos preliminares y objetivos del estudio. La integración de VOS fue consistente entre los pacientes y solo el 13,3 % de ellos sufrió una recurrencia de la infección por C. difficile (CDI) en un amplio rango de dosis administradas en un estudio de fase I abierto. Sin embargo, VOS no alcanzó su criterio de valoración principal de eficacia en un estudio de fase II aleatorizado y controlado con placebo de seguimiento, en el que se administró a una dosis fija que había demostrado eficacia en la fase I. En respuesta a ello, este grupo llevó a cabo varios estudios de metabolómica de seguimiento. El objetivo de su primer estudio de seguimiento fue conciliar estos resultados dispares mediante la evaluación de los cambios inducidos por el VOS en el microbioma y en los metabolitos asociados a los microbios [3]. Su segundo objetivo se describe en la siguiente sección titulada «Éxito en un ensayo de fase III a pesar del fracaso en la fase II».
Métodos. En los ensayos clínicos de fase I y II, se evaluó la colonización de VOS comparando el número de especies bacterianas asociadas a VOS en muestras de heces recogidas 1, 4 y 8 semanas después de la administración, y la recurrencia de la CDI se determinó en función de los síntomas clínicos. Se llevaron a cabo análisis metagenómicos completos mediante secuenciación «shotgun» y análisis metabolómicos globales de forma retrospectiva sobre los conjuntos de datos de las fases I y II para evaluar el impacto de VOS en los resultados clínicos.
Resultados. En el ensayo clínico de fase II, los participantes tratados que no desarrollaron una CDI recurrente presentaban un número significativamente mayor de especies de VOS una semana después de la administración del tratamiento que aquellos que sufrieron una recurrencia. Se llevó a cabo un análisis post hoc de los resultados de los ensayos de fase I y fase II para evaluar el grado de injerto una semana después de que los participantes recibieran una dosis alta (fase I), baja (fase I) o fija (fase II). Los sujetos de la fase 1 que recibieron la dosis alta presentaban un número significativamente mayor de especies de VOS en la semana 1 que las otras dos cohortes de dosificación (Figura 7). Además, se observó que el injerto en la semana 1 era muy variable en los participantes de la fase 2, lo que sugiere que el éxito del injerto dependía de la dosis y que la dosificación fue subóptima en el ensayo de fase 2.
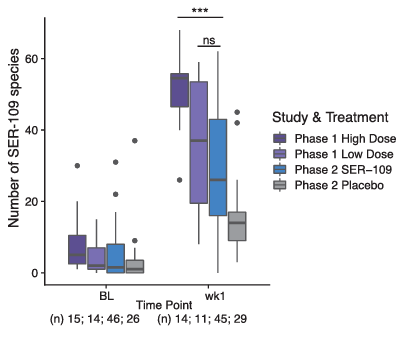
Figura 7. Relación entre el injerto de especies de VOS y las dosis administradas en los estudios de fase 1 y 2. Los participantes que recibieron la dosis alta en el estudio de fase 1 presentaban un número significativamente mayor de especies de VOS que los participantes del grupo de tratamiento del estudio de fase 2 que recibieron una dosis fija baja (p < 0,001). Imagen reproducida de McGovern et al., Clin Infect Dis, 2021, con licencia CC BY 4.0.
Curiosamente, también se observó una correlación positiva significativa entre el número de especies de VOS y la abundancia de los ácidos biliares secundarios, el ácido litocólico (LCA) y el ácido desoxicólico (DCA), lo que sugiere que un mayor asentamiento de especies conduce a una mayor conversión de los ácidos biliares primarios conjugados en ácidos biliares secundarios, un paso clave para frenar el crecimiento de C. difficile y recuperar la salud intestinal normal (Figura 8).
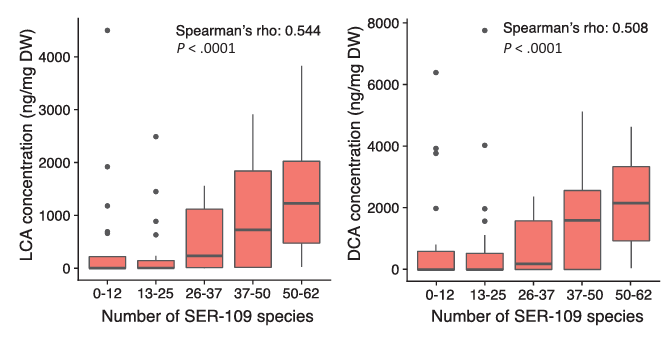
Figura 8. Relación entre el injerto de especies de VOS (anteriormente SER-109) y la concentración de ácidos biliares secundarios en el estudio de fase II. El número de especies de VOS una semana después de la administración se asoció significativamente con la concentración de LCA y DCA. Abreviaturas: DCA = ácido desoxicólico, DW = peso seco, LCA = ácido litocólico. Imagen reproducida de McGovern et al., Clin Infect Dis, 2021, con licencia CC BY 4.0.
Conclusiones del estudio
- Este estudio demostró que el ensayo clínico de fase II probablemente fracasó debido a una dosificación subóptima que provocó un injerto irregular, y no por una eficacia terapéutica insuficiente.
- Estos hallazgos también dieron lugar a la hipótesis de que el injerto de VOS provoca cambios funcionales importantes a través de metabolitos clave asociados a los microbios, lo que rompe el ciclo de la CDI recurrente.
- En este primer estudio de seguimiento, la metabolómica resultó fundamental para demostrar por qué los resultados de los ensayos de fase I y fase II no coincidían entre sí, lo que permitió al equipo seguir adelante con un ensayo de fase III rediseñado a pesar del fracaso en la fase II, evitando así que una terapia prometedora quedara archivada y salvando la inversión comercial. El siguiente estudio de caso analiza los resultados del ensayo de fase III de este grupo.
Un ensayo de fase III satisfactorio a pesar del fracaso en la fase II
Datos preliminares y objetivos del estudio. A la luz de los resultados descritos en el estudio de caso anterior, los investigadores rediseñaron el estudio de fase III para aplicar una estrategia de dosificación más eficaz [4]. A los participantes del grupo de tratamiento se les administró una dosis fija de VOS diez veces superior a la administrada en la fase II, con el objetivo de evaluar la eficacia terapéutica del VOS en condiciones adecuadas para el injerto.
Métodos. La eficacia se evaluó en función de la reducción del riesgo relativo y de la respuesta clínica sostenida hasta 8 semanas después de la administración. Concretamente, el riesgo relativo se definió como el porcentaje de pacientes con recidiva en el grupo de VOS dividido por el porcentaje en el grupo de placebo. La respuesta clínica sostenida se definió como la ausencia de recidiva durante las 8 semanas de seguimiento de los pacientes. Se realizaron análisis metabolómicos globales en muestras de heces recogidas 1, 2 y 8 semanas después de la administración para evaluar los perfiles de ácidos biliares en el intestino.
Resultados. Se observó que el VOS (anteriormente denominado SER-109) era superior al placebo a la hora de reducir el riesgo de recurrencia de la infección por C. difficile y de generar una respuesta clínica sostenida en los pacientes (Figura 9).
El grupo tratado con VOS mostró un aumento significativamente mayor de los ácidos biliares secundarios con respecto al valor basal, en comparación con el grupo de placebo (Figura 10), lo que coincidió con una reducción de las enterobacterias proinflamatorias y un aumento de las bacterias del género Firmicutes, especies que se consideran supresores reguladores clave de la infección por Clostridioides difficile (CDI).
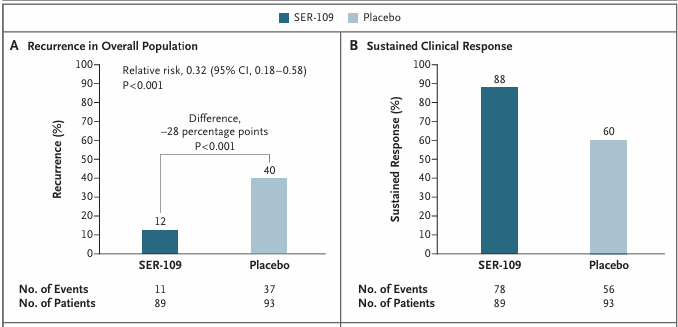
Figura 9. Recurrencia de la CDI hasta las 8 semanas. (A) El análisis de eficacia primario demostró la superioridad de VOS (SER-109) frente al placebo en la reducción del riesgo de recurrencia de la CDI. (B) VOS también fue superior al placebo en la consecución de una respuesta clínica sostenida. Imagen reproducida de Feuerstadt et al., N Engl J Med, 2022, bajo licencia CC BY 4.0.
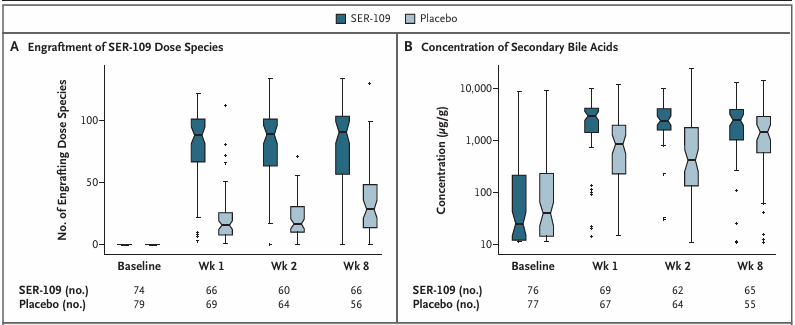
Figura 10. Cambios composicionales y metabolómicos en el microbioma tras recibir VOS (SER-109) o placebo. (A) El número de especies que se han implantado y (B) la concentración de ácidos biliares secundarios combinados detectados en las muestras de heces de los pacientes 1, 2 y 8 semanas después de recibir VOS o placebo. Imagen reproducida de Feuerstadt et al., N Engl J Med, 2022, con licencia CC BY 4.0.
Conclusiones del estudio
- Un ensayo de fase III rediseñado para optimizar la dosificación a partir de datos metabolómicos permitió que el VOS se implantara correctamente y, por lo tanto, demostrara su superioridad frente al placebo en la prevención de la recurrencia de la infección por Clostridioides difficile.
- Los datos metabolómicos de este estudio y de los estudios clínicos anteriores sugieren que la integración de las VOS depende de la dosis, y que su mecanismo de acción podría estar relacionado con la producción de ácidos biliares secundarios.
- Estos hallazgos dieron lugar a una hipótesis muy plausible sobre el mecanismo de acción del VOS. Dado que comprender el mecanismo de acción de un agente terapéutico es esencial para optimizar la eficacia del tratamiento, predecir los efectos secundarios y determinar las dosis adecuadas, este estudio podría facilitar la toma de decisiones rentables para los estudios clínicos de seguimiento.
Descubrimiento de biomarcadores diagnósticos de alto rendimiento que pasan desapercibidos para otras ciencias ómicas
Introducción. Las tasas de supervivencia del cáncer colorrectal (CCR) podrían mejorarse mediante la detección precoz, por lo que existe una gran necesidad de biomarcadores no invasivos que faciliten esta tarea. Los cambios en el metabolismo energético son un rasgo característico de la tumorigénesis, ya que las células cancerosas dependen de la glucólisis aeróbica para generar energía e intermedios glucolíticos, que sirven como precursores de lípidos y aminoácidos. Las diferencias en los perfiles de aminoácidos y lípidos en plasma y suero han demostrado una alta sensibilidad y especificidad a la hora de distinguir a los pacientes con CCR de los controles sanos.
Datos preliminares y objetivos del estudio. Cada vez hay más pruebas que sugieren que las plaquetas contribuyen a la progresión del cáncer. Los experimentos de cocultivo han demostrado que el plasma rico en plaquetas (PRP) favorece un fenotipo proliferativo en las células cancerosas, y se cree que la adhesión de las plaquetas facilita la metástasis tumoral. En este estudio, los investigadores plantearon la hipótesis de que el PRP con alta actividad biológica podría revelar cambios metabólicos que ayudaran a la detección precoz del CCR. El objetivo de este estudio fue analizar los perfiles metabólicos del plasma rico en plaquetas (PRP) en pacientes con CCR y controles sanos para identificar cambios metabólicos que pudieran ayudar a detectar el CCR en fase temprana [5].
Métodos. Se llevó a cabo un análisis metabolómico global del PRP recogido y agrupado en tres grupos: pacientes con CCR en estadio I y II, pacientes con CCR en estadio III y IV, y controles sanos. Se utilizaron el análisis de conglomerados y el enriquecimiento de clases para evaluar las diferencias en los perfiles entre las tres cohortes.
Resultados. Los niveles de lactosa, glicerol-3-fosfato, ácido 2-hidroxiglutárico, ácido isocítrico y ácido cítrico resultaron significativamente elevados en el PRP extraído de pacientes con CCR en comparación con los controles (Figura 11). Aunque los niveles de estos metabolitos no mostraron diferencias significativas entre los tumores en estadio temprano (estadios I y II) y los de estadio avanzado (estadios III y IV), sus concentraciones sistemáticamente más elevadas en comparación con los controles no descartaban su uso como biomarcadores candidatos para la detección precoz del CCR.
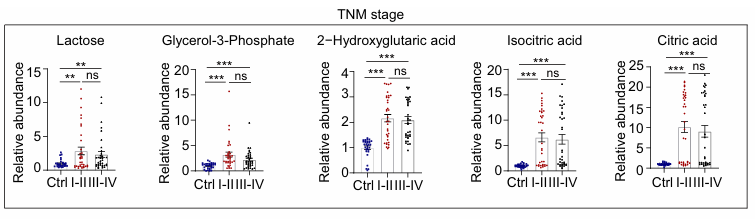
Figura 11. Abundancia relativa de cinco metabolitos clave de los hidratos de carbono en diferentes estadios de la progresión del cáncer colorrectal. Se observaron diferencias significativas en estos metabolitos entre los pacientes con cáncer colorrectal y los controles. No se observaron diferencias entre los tumores en estadio temprano (estadios I y II) y los de estadio avanzado (estadios III y IV). Imagen reproducida de Hu et al., Mol Oncol, 2025, bajo licencia CC BY 4.0.
Se comparó el rendimiento diagnóstico de esta firma de 5 metabolitos con el del antígeno carcinoembrionario (CEA) y el antígeno carbohidrato 19-9 (CA 19-9), dos biomarcadores sanguíneos clásicos del cáncer colorrectal. El CEA mostró una sensibilidad del 80 %, una especificidad del 76,67 % y un valor del área bajo la curva ROC (AUC) de 0,783; el CA 19-9 alcanzó una sensibilidad del 74 %, una especificidad del 63 % y un AUC de 0,641. Por el contrario, la firma metabolómica obtuvo unos resultados mucho mejores, distinguiendo el CCR de los controles sanos con una sensibilidad del 90 %, una especificidad del 96,67 % y un AUC de 0,961. (Figura 12).
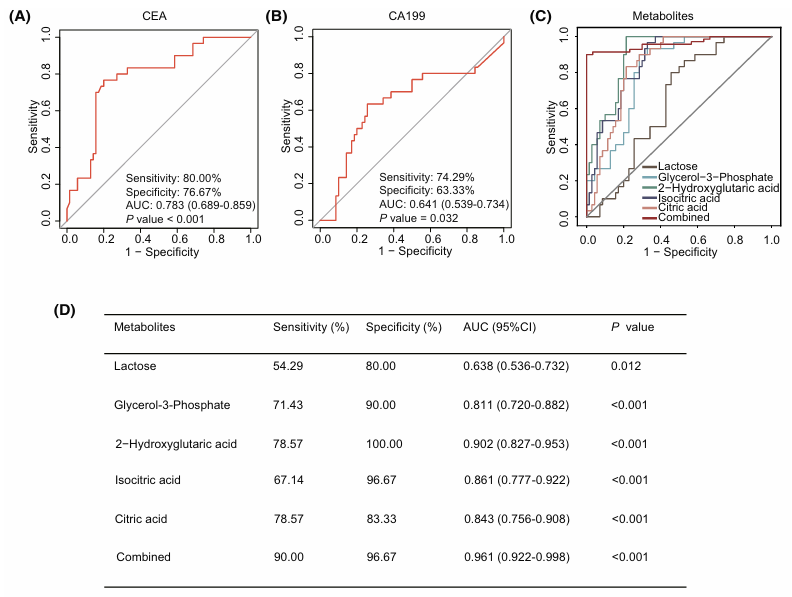
Figura 12. Rendimiento diagnóstico de la firma metabólica frente a los biomarcadores tradicionales del CCR. (A) Eficacia diagnóstica del CEA, (B) del CA 19-9 y (C) de cada metabolito de la firma, tanto por separado como en combinación. (D) Sensibilidad, especificidad y área bajo la curva (AUC) de cada metabolito de la firma a la hora de distinguir a los pacientes con CCR de los controles sanos. Imagen reproducida de Hu et al., Mol Oncol, 2025, con licencia CC BY 4.0.
Nutrición humana
Identificación de los vínculos mecánicos entre la alimentación y la enfermedad renal crónica
Introducción. La enfermedad renal crónica (ERC) afecta a 800 millones de personas en todo el mundo y supone un coste anual de aproximadamente 407 000 millones de dólares. La alimentación influye de manera significativa en la salud renal y, según hallazgos recientes, los patrones alimentarios basados en vegetales podrían ofrecer una forma rentable de prevenir la ERC. El Estudio de Salud Adventista 2 (AHS-2) es un estudio plurianual destinado a evaluar las relaciones entre el estilo de vida, la alimentación y la enfermedad. Hallazgos recientes de esta cohorte mostraron una asociación entre los patrones alimentarios vegetarianos y una menor incidencia de diabetes e hipertensión, dos factores de riesgo clave para la ERC. La dieta vegetariana también se asoció fuertemente con una menor mortalidad por enfermedad renal. A pesar de estas fuertes asociaciones, no estaba claro cómo los patrones alimentarios vegetarianos parecían proteger la salud renal.
Datos preliminares y objetivos del estudio. Análisis metabolómicos previos realizados en la cohorte ASH-2 revelaron que las personas veganas presentaban niveles plasmáticos más elevados de metabolitos derivados de polifenoles, de origen microbiano y relacionados con las vitaminas, mientras que las personas no vegetarianas presentaban niveles más elevados de lípidos y aminoácidos proinflamatorios. Con el objetivo de comprender mejor las posibles relaciones de causa y efecto entre el riesgo de ERC, su progresión y la dieta, este estudio examinó las asociaciones entre los patrones alimentarios vegetarianos, la función renal y los metabolitos relacionados con la dieta utilizando la tasa de filtración glomerular estimada (eGFR) como medida de la salud renal [6].
Métodos. Se agrupó a 899 participantes de la cohorte AHS-2 según sus patrones alimentarios, que incluían dietas veganas, pesco-vegetarianas y no vegetarianas. Se llevó a cabo un análisis metabolómico global de las muestras de suero recogidas de estos participantes en el estudio. Se utilizaron los niveles de creatinina sérica para calcular la TFGe. Para el análisis de las subclases de metabolitos, se promediaron los valores de los metabolitos componentes individuales dentro de una subclase para obtener una media compuesta para dicha subclase. Las puntuaciones T se obtuvieron dividiendo la desviación estándar de las medias compuestas, teniendo en cuenta las covarianzas entre los metabolitos. Se determinó el número de metabolitos significativos o diferenciales para cada subclase, junto con el número de metabolitos diferenciales asociados positiva o inversamente con el patrón alimentario.
Resultados. Numerosos metabolitos se asociaron de forma exclusiva con la dieta y los niveles de creatinina, lo que reveló conexiones específicas entre los patrones alimentarios y el fenotipo metabólico. Por ejemplo, en comparación con las personas no vegetarianas, los veganos presentaban niveles significativamente más bajos de ciertas toxinas urémicas, como el p-cresol, el sulfato de 3-metilcatecol y el glucurónido, que se asocian con una TFG estimada (eGFR) más baja y, por consiguiente, con una función renal más reducida. Análisis posteriores identificaron subclases de metabolitos diferenciales (en lugar de metabolitos individuales) asociadas con diferentes patrones alimentarios y la creatinina sérica. Las comparaciones entre los patrones alimentarios veganos y no vegetarianos mostraron que los plasmalógenos y los lisoplasmalógenos tenían la mayor proporción de metabolitos significativos en uno o ambos grupos (Figura 13). Se ha demostrado que los plasmalógenos y los lisoplasmalógenos median la reparación de la membrana durante el estrés oxidativo. Se han observado niveles circulantes reducidos de estas subclases de metabolitos en pacientes con ERC avanzada, probablemente debido a un elevado estrés oxidativo.
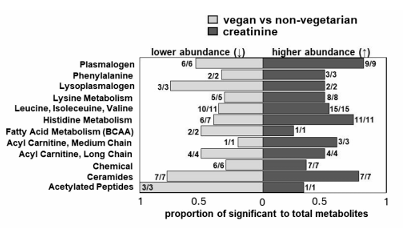
Figura 13. Subclases diferenciales que muestran asociaciones opuestas entre los patrones alimentarios y la creatinina sérica con los metabolitos plasmáticos. Las subclases con abundancia diferencial se determinaron tras calcular la media compuesta (promediando los metabolitos de los componentes) para cada grupo alimentario y, posteriormente, las puntuaciones t. Las barras reflejan la proporción de metabolitos diferenciales respecto al total de metabolitos del panel asociados positivamente con la creatinina (a la derecha del 0) o inversamente con el patrón alimentario indicado (a la izquierda del 0). Las ratios representan el número de metabolitos significativos asociados en la misma dirección (inversamente o positivamente) respecto al número total de metabolitos significativos en la subclase. La dirección predominante del cambio en la abundancia de cada subclase, que refleja la dirección general de los metabolitos componentes, se representa gráficamente para los contrastes de creatinina entre veganos y no vegetarianos. Imagen reproducida de Butler et al., J Ren Nutr, 2025, con licencia CC BY 4.0.
Conclusiones del estudio
- Este estudio identificó cambios metabólicos propios de distintos patrones alimentarios, con el fin de revelar los vínculos mecánicos entre la alimentación y la TFGe.
- La metabolómica permitió a los investigadores ir más allá de la medición habitual de la creatinina para detectar los cambios metabólicos subyacentes, lo que aportó pruebas más sólidas de que las vías metabólicas determinadas por la dieta están directamente relacionadas con la salud renal.
Superar las limitaciones de los informes alimentarios autoinformados mediante la estandarización de las huellas biomarcadoras de la ingesta de alimentos
Introducción. Los métodos tradicionales de evaluación de la dieta, que a menudo se basan en la información facilitada por los propios participantes, son propensos a errores y sesgos. La metabolómica puede subsanar esta limitación al detectar los compuestos derivados de los alimentos, lo que proporciona una huella molecular objetiva de la ingesta alimentaria. Aunque se han identificado numerosos biomarcadores dietéticos, la mayoría de los estudios se han centrado en alimentos concretos en lugar de en patrones alimentarios completos, y son pocos los ensayos controlados de alimentación que han incluido un diseño cruzado aleatorio para reducir la heterogeneidad a la hora de cuantificar las asociaciones con dietas específicas.
Datos preliminares y objetivos del estudio. Con el fin de subsanar las lagunas existentes en los informes metabolómicos actuales sobre la alimentación, este estudio llevó a cabo un ensayo aleatorio cruzado y controlado en el que se comparó una dieta australiana saludable (HAD), basada en las Directrices dietéticas australianas vigentes, con una dieta australiana típica (TAD). Los objetivos de los investigadores eran: 1) describir los cambios metabólicos en plasma y orina en respuesta a estos dos patrones dietéticos, y 2) identificar metabolitos discriminatorios específicos de la dieta saludable con el fin de desarrollar una puntuación de calidad de la dieta derivada del metaboloma que pudiera ayudar a evaluar la asociación de la dieta saludable con marcadores de salud cardiometabólica [7].
Métodos. En un ensayo alimentario aleatorizado cruzado de ocho semanas de duración, adultos sanos completaron dos intervenciones dietéticas controladas de dos semanas de duración —una dieta saludable y una dieta habitual—, separadas por un periodo de lavado. Se proporcionaron todas las comidas para garantizar el cumplimiento y minimizar la variabilidad de la dieta. Se recogieron muestras de sangre y orina en ayunas antes y después de cada intervención, que luego se analizaron mediante perfiles metabolómicos no dirigidos. Se utilizó la regresión de red elástica para identificar los metabolitos que mejor distinguían la dieta saludable de la dieta habitual. Estos metabolitos se utilizaron para construir una puntuación compuesta de la calidad de la dieta derivada del metaboloma, que luego se evaluó para detectar asociaciones con marcadores cardiometabólicos, incluyendo la presión arterial, los lípidos y la glucosa en ayunas.
Resultados. Tras dos semanas, se observaron diferencias con respecto a los valores iniciales en 408 metabolitos plasmáticos y 319 metabolitos urinarios, muchos de los cuales pertenecían a las vías metabólicas de los lípidos, los aminoácidos y los xenobióticos. La comparación directa de las dos dietas permitió identificar cientos de metabolitos que presentaban diferencias significativas. Cuarenta y tres de ellos mostraron cambios consistentes tanto en plasma como en orina, lo que sugiere que cada patrón dietético generaba una firma metabólica sólida (Figura 14). El análisis de aprendizaje automático identificó 31 metabolitos plasmáticos y 34 urinarios que distinguían mejor la dieta saludable (HAD) de la dieta típica (TAD). Estos metabolitos se utilizaron para crear la puntuación de calidad de la dieta derivada del metaboloma (Figura 15 A, B). Las puntuaciones más altas, que indicaban una estrecha adherencia a la HAD, se asociaron significativamente con una mejora de los marcadores cardiometabólicos, incluyendo una menor presión arterial, triglicéridos, colesterol LDL y glucosa plasmática en ayunas (Figura 15 C-H).
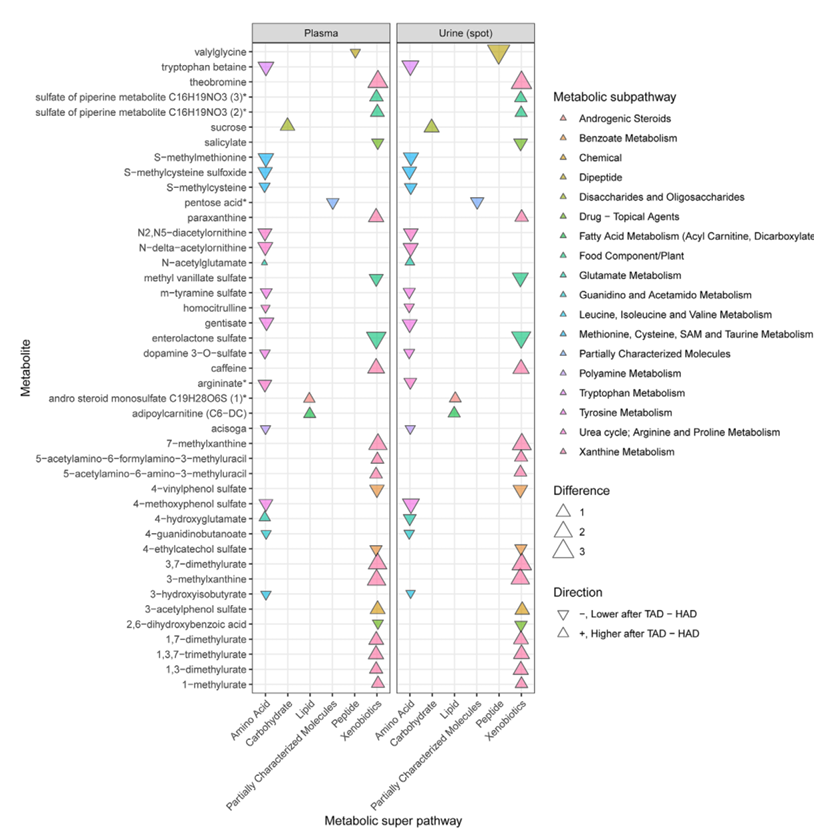
Figura 14. Magnitud y dirección del cambio en 43 metabolitos tras dos semanas de seguir una dieta HAD en comparación con una dieta TAD. Las concentraciones de estos metabolitos mostraron diferencias sistemáticas entre los grupos, tanto en plasma como en orina. Imagen reproducida de Stanford et al., Mol Nutr Food Res, 2025, bajo licencia CC BY 4.0.
Figura 15. Desarrollo de una puntuación de biomarcadores de la calidad de la dieta derivada del metaboloma y su asociación con marcadores de salud cardiometabólica. Se muestran los metabolitos discriminativos y sus coeficientes en (A) plasma y (B) orina. Los coeficientes se utilizaron para construir una puntuación compuesta de la calidad de la dieta, en la que una puntuación más alta se asoció con una mejora significativa de los resultados cardiometabólicos, incluyendo (C) una presión arterial sistólica más baja, (D) la presión arterial diastólica, (E) los triglicéridos, (F) el colesterol LDL, (G) el colesterol HDL y (H) la glucosa plasmática en ayunas. Imagen reproducida de Stanford et al., Mol Nutr Food Res, 2025, con licencia CC BY 4.0.
Conclusiones del estudio
- En este estudio, los investigadores utilizaron la metabolómica para obtener una puntuación que mide cuantitativamente el cumplimiento de la dieta, lo que permite evitar los sesgos y errores inherentes a la información sobre la ingesta alimentaria facilitada por los propios participantes. De cara al futuro, esta puntuación de la calidad de la dieta podría mejorar significativamente la interpretabilidad y la reproducibilidad de los estudios sobre nutrición, lo que a su vez permitiría mejorar la eficiencia y la rentabilidad de los diseños de estudios futuros.
Desarrollo de herramientas para mejorar la cuantificación del consumo de alimentos ultraprocesados
Introducción. Los alimentos ultraprocesados (AUP) se caracterizan por contener ingredientes y aditivos industriales que no suelen utilizarse en la cocina casera, como edulcorantes artificiales y almidones modificados. Aunque el procesamiento de los alimentos desempeña un papel importante en la seguridad alimentaria y la vida útil de los productos, los estudios observacionales han asociado de forma sistemática un elevado consumo de AUP con un mayor riesgo de obesidad, enfermedades cardiovasculares, cáncer y mortalidad por todas las causas. En la actualidad, los estudios nutricionales sobre los UPF se ven limitados por la falta de herramientas que permitan medir con precisión su ingesta. Los cuestionarios habituales de evaluación de la dieta y los recordatorios de 24 horas carecen de la información detallada sobre ingredientes y procesamiento necesaria para clasificar los alimentos según los sistemas de clasificación de la industria.
Datos preliminares y objetivos del estudio. El objetivo de este estudio era abordar la necesidad de contar con biomarcadores objetivos que permitan evaluar de forma fiable el consumo de alimentos procesados ultraprocesados (UPF) y sus efectos metabólicos, con el objetivo a largo plazo de mejorar el estudio de las relaciones entre la alimentación y las enfermedades [8].
Métodos. Los ensayos de alimentación controlada, en los que se proporcionan todos los alimentos y se supervisa de cerca su ingesta, son ideales para identificar biomarcadores relevantes de los alimentos procesados ultrapreparados (UPF). Este estudio fue un análisis secundario de un ensayo aleatorizado, cruzado y de alimentación controlada en el que 20 adultos consumieron una dieta rica en UPF o una dieta totalmente exenta de UPF. Las dietas se igualaron en cuanto a calorías y nutrientes. Se recogieron muestras de sangre y orina en múltiples momentos a lo largo del estudio y se analizaron mediante perfiles metabolómicos globales. Se compararon los niveles de metabolitos entre las dietas utilizando modelos lineales de efectos mixtos, y se realizaron análisis adicionales para evaluar los cambios a nivel de las vías metabólicas.
Resultados. En comparación con la dieta de control, la dieta rica en alimentos procesados de alta densidad (UPF) indujo cambios metabólicos generalizados y consistentes. Tal y como se esperaba, varios metabolitos directamente relacionados con los aditivos y conservantes alimentarios aumentaron de forma significativa en la dieta rica en UPF, mientras que los metabolitos asociados a las frutas y verduras enteras se redujeron, lo que refleja una menor ingesta de estos alimentos. También se observaron cambios en las vías relacionadas con el metabolismo lipídico, los ácidos biliares, los aminoácidos y los metabolitos de la microbiota intestinal, lo que demuestra que el consumo de UPF altera tanto el metabolismo del huésped como el relacionado con la microbiota. Es importante destacar que muchos de estos cambios en los metabolitos fueron consistentes tanto en plasma como en orina, lo que respalda su potencial como biomarcadores sólidos de la ingesta de UPF y la respuesta metabólica a corto plazo (Figura 16).
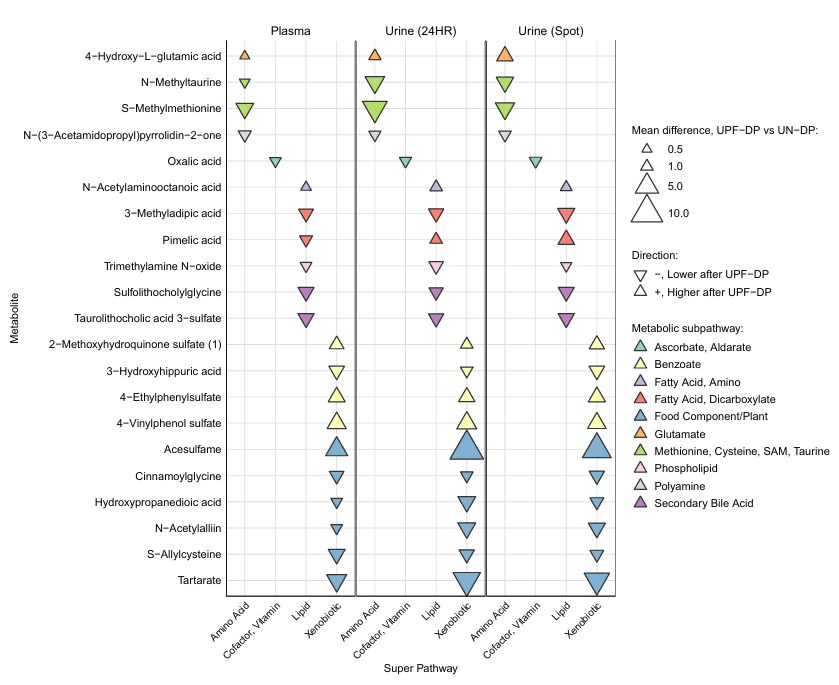
Figura 16. Magnitud y dirección de las diferencias medias en la semana 2 en los metabolitos tras el consumo de una dieta rica en alimentos ultraprocesados (UPF) en comparación con una dieta de control. Los metabolitos enumerados aquí (n = 21) mostraron diferencias entre los participantes individuales (n = 20) entre los grupos UPF-DP y UN-DP en la semana 2 para el plasma, así como en la semana 1 y la semana 2 para la orina de 24 horas. Las diferencias medias en los metabolitos se estimaron mediante un modelo lineal mixto ajustado por dieta, fase, secuencia y momento temporal solo para la orina, y se calculó la ingesta energética (EI) durante la semana previa a la recogida de muestras con intersecciones aleatorias específicas para cada sujeto. Imagen reproducida de O'Connor et al., J Nutr, 2023, con licencia CC BY 4.0.
Conclusiones del estudio
- Este estudio demuestra que seguir una dieta rica en alimentos ultraprocesados (UPF) provoca cambios claros y cuantificables en el metaboloma plasmático y urinario. Los metabolitos directamente relacionados con los UPF —como los conservantes de benzoato y los edulcorantes artificiales—, así como aquellos asociados a alteraciones en la microbiota, podrían servir como biomarcadores objetivos del consumo de UPF.
- Estos biomarcadores podrían ayudar a superar algunas limitaciones que desde hace tiempo presentan los métodos tradicionales de evaluación de la dieta y mejorar la precisión y la reproducibilidad de los estudios destinados a caracterizar la relación entre la alimentación y la salud.
Salud de las mascotas y cría de animales
Caracterización del mecanismo de acción de un tratamiento muy utilizado pero poco conocido para la diarrea crónica del intestino grueso en perros
Introducción. La diarrea crónica del intestino grueso en perros es un trastorno gastrointestinal frecuente provocado por interacciones complejas entre la microbiota intestinal, la dieta, el metabolismo del huésped, las respuestas inmunitarias y los factores ambientales. Las pruebas recientes sugieren que la disbiosis microbiana contribuye a esta afección al alterar los productos metabólicos en el intestino, incluida la producción de metabolitos inflamatorios o nocivos. La suplementación de la dieta con fibra puede mejorar la salud gastrointestinal al favorecer la fermentación microbiana beneficiosa y aumentar la producción de postbióticos protectores, como los ácidos grasos de cadena corta (AGCC), los ácidos biliares secundarios y los indoles. Aunque la intervención nutricional se considera el tratamiento principal para la diarrea crónica del intestino grueso en perros, los mecanismos que subyacen a su efecto terapéutico son poco conocidos.
Datos preliminares y objetivos del estudio. Investigadores de Hill’s Pet Nutrition, Inc. demostraron que una mezcla patentada de fibras solubles e insolubles, seleccionadas por su actividad prebiótica y postbiótica, mejoraba los parámetros de las heces. El objetivo de este estudio de seguimiento era evaluar el impacto mecánico de una intervención dietética con suplementos de fibra sobre la salud gastrointestinal en perros que padecían diarrea del intestino grueso [9].
Métodos. Los investigadores llevaron a cabo un ensayo clínico de ocho semanas de duración con perros adultos que padecían diarrea crónica del intestino grueso. Todos los perros recibieron una dieta terapéutica rica en fibra, formulada con múltiples fibras vegetales solubles e insolubles. Se recogieron muestras fecales y séricas al inicio del estudio y en varios momentos durante la intervención. Se utilizó la metabolómica no dirigida para analizar las muestras fecales y séricas con el fin de evaluar los cambios en la salud gastrointestinal y la actividad metabólica.
Resultados. La intervención dietética con suplementos de fibra mejoró rápidamente los signos clínicos de la diarrea. Se observaron aumentos significativos en la consistencia de las heces en el plazo de un día y la diarrea se resolvió en la mayoría de los perros en el plazo de varias semanas. Los análisis metabolómicos revelaron cambios significativos en los metabolitos fecales y circulantes que indicaban una transición en el metabolismo microbiano intestinal, pasando de la fermentación de proteínas (putrefacción) a la fermentación de carbohidratos (sacarólisis) (Figura 17). La intervención redujo los metabolitos asociados a la degradación bacteriana de proteínas y aumentó los metabolitos de origen vegetal y los postbióticos microbianos beneficiosos, incluidos los derivados del indol. Otros cambios metabolómicos sugirieron mejoras en las vías inflamatorias, el metabolismo lipídico, la integridad de la barrera intestinal y las interacciones entre el huésped y los microbios.
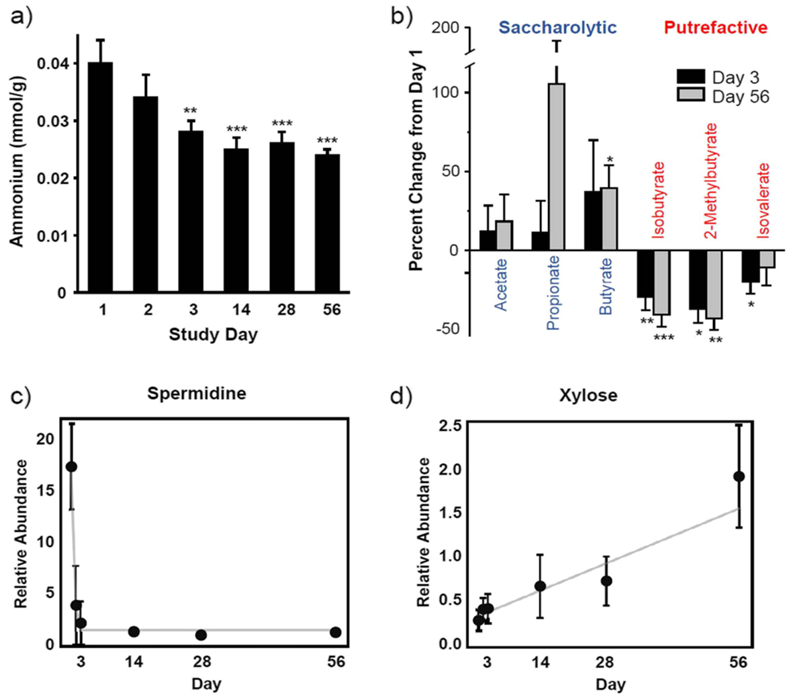
Figura 17. Efecto de la intervención dietética sobre el metabolismo fermentativo. La intervención con fibra dietética modificó el metabolismo, pasando de la fermentación de proteínas al metabolismo de los hidratos de carbono, como lo demuestran las reducciones en (A) las concentraciones fecales de amonio (A), los ácidos grasos de cadena corta (B) y la espermidina (C), así como el aumento del monosacárido xilosa (D). Imagen reproducida de Fritsch et al., BMC Vet Res, 2022, con licencia CC BY 4.0.
Conclusiones del estudio
- En general, los resultados de este estudio indican que la suplementación con fibra dietética modifica positivamente tanto el metabolismo microbiano como la fisiología gastrointestinal del huésped, lo que demuestra cómo la fibra dietética ejerce su efecto terapéutico.
- La metabolómica permitió a los investigadores medir directamente los productos metabólicos generados en el intestino, lo que proporcionó una visión funcional de estos mecanismos que no se habría podido deducir de otro modo.
- Los datos metabolómicos aportaron pruebas mecánicas de la eficacia de la intervención nutricional, que podrían utilizarse para respaldar las declaraciones de marketing.
Orientaciones sobre la nutrición en las primeras etapas de vida para reducir el sacrificio de vacas lecheras
Introducción. La nutrición durante las primeras etapas de la vida desempeña un papel crucial en el desarrollo de la salud metabólica a largo plazo. En el caso de los terneros lecheros, una nutrición insuficiente antes del destete se asocia con un crecimiento reducido, una función inmunitaria deteriorada y una menor productividad a largo plazo, mientras que una nutrición adecuada tiende a aumentar la producción de leche y favorece la madurez reproductiva temprana. Los mecanismos biológicos que relacionan la nutrición durante las primeras etapas de la vida con la productividad en las vacas adultas están bien documentados, pero aún se conocen poco.
Datos preliminares y objetivos del estudio. Estudios previos han demostrado que un aporte adecuado de nutrientes antes del destete modifica las vías metabólicas relacionadas con la energía, el metabolismo de los aminoácidos y la sensibilidad a la insulina, cambios que persisten en la edad adulta. En este estudio, los investigadores plantearon la hipótesis de que un mayor aporte de nutrientes antes del destete mejora el metabolismo energético y la productividad a largo plazo en las vacas lecheras [10]. El objetivo de este estudio fue poner a prueba su hipótesis determinando cómo la ingesta de leche en las primeras etapas de la vida influye en los perfiles metabolómicos, la producción de leche y la supervivencia a lo largo de múltiples lactancias.
Métodos. Se estudió a 78 vacas Holstein desde el primer parto hasta el quinto. Los terneros se asignaron aleatoriamente a un grupo con ración elevada en nutrientes, que recibió 8 litros de sustituto de leche al día, o a un grupo con ración restringida, que recibió 4 litros al día hasta el destete. Se monitorizaron el crecimiento, el rendimiento reproductivo, la producción de leche, el consumo de pienso y la supervivencia a lo largo de las lactancias. Se recogieron muestras de suero el día 60 de la lactancia y se analizaron mediante metabolómica no dirigida para evaluar el estado metabólico durante el pico de producción de leche.
Resultados. Los terneros que recibieron niveles elevados de sustituto de leche mostraron un mayor crecimiento inicial y una mejor sensibilidad a la insulina en fases posteriores de su desarrollo, en comparación con los terneros con alimentación restringida. Las vacas alimentadas con niveles elevados de sustituto de leche también produjeron mayores rendimientos de grasa láctea y un mayor contenido de grasa, y produjeron más leche cuando los volúmenes se corrigieron en función de los porcentajes de grasa corporal y proteína. El rendimiento reproductivo fue similar entre los grupos en la primera lactancia, pero en la segunda lactancia las vacas alimentadas con un sustituto lácteo elevado tuvieron una tasa de concepción más alta tras el primer apareamiento que las vacas con alimentación restringida. El análisis de supervivencia mostró que las vacas que recibieron una nutrición más elevada en las primeras etapas de vida tenían un riesgo sustancialmente menor de ser sacrificadas antes de los partos posteriores (Figura 18). El perfil metabolómico en el pico de la lactancia reveló diferencias metabólicas persistentes entre los grupos, incluyendo cambios en los metabolitos de aminoácidos, lípidos, nucleótidos y carbohidratos (Figura 19). Los análisis de las vías metabólicas identificaron alteraciones en el metabolismo de las purinas, las pirimidinas, la esfingosina, los ácidos grasos y el ciclo de Krebs, lo que sugiere que la nutrición en las primeras etapas de vida influye en la programación metabólica a largo plazo.
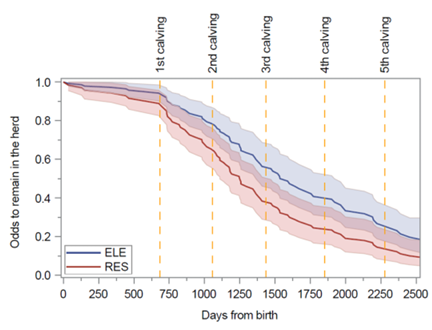
Figura 18. Probabilidad de que las vacas permanezcan en el rebaño, y por lo tanto eviten el sacrificio, en función de los días transcurridos desde el nacimiento para las vacas alimentadas con una ingesta nutricional aumentada (línea azul) o restringida (línea roja) antes del destete. Imagen reproducida de Leal et al., J Dairy Sci, 2025, bajo licencia CC BY 4.0.
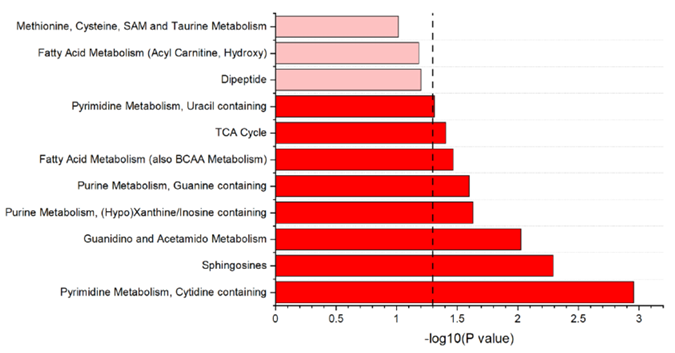
Figura 19. Visualización de las vías metabólicas significativas puestas de manifiesto por el análisis de enriquecimiento basado en los metabolitos detectados en el suero de vacas lecheras en el pico de la lactancia. Las vías metabólicas representadas en rojo más oscuro tenían un valor de p < 0,05; las representadas en rojo más claro tenían un valor de p < 0,10. Imagen reproducida de Leal et al., J Dairy Sci, 2025, bajo licencia CC BY 4.0.
Conclusiones del estudio
- Los resultados de este estudio demuestran que existe una relación directa entre la nutrición durante las primeras etapas de la vida y la función metabólica a largo plazo, lo que puede servir de base para tomar decisiones sobre la alimentación que, en última instancia, mejoren la productividad y la sostenibilidad a lo largo de toda la vida del ganado lechero, lo que se traduce en un ahorro de costes y en beneficios nutricionales para las personas.
Mejora de la calidad de la carne de vacuno mediante la caracterización de la relación entre la calidad del suelo, la alimentación y la salud del ganado
Introducción. En la industria cárnica estadounidense, el ganado suele alimentarse con dietas a base de maíz durante los últimos meses de vida para maximizar su crecimiento y la calidad de la canal. La producción de cultivos forrajeros depende de monocultivos, fertilizantes sintéticos y riego, lo que agota los nutrientes del suelo, degrada su estructura y contamina las fuentes de agua. Se ha demostrado que el engorde de los animales en pastos mediante prácticas de pastoreo adaptativo y rotativo mejora la diversidad vegetal, el carbono del suelo y el funcionamiento general del ecosistema. Los animales que pastan forrajes frescos consumen mayores cantidades y una mayor variedad de polifenoles, y el enriquecimiento de estos compuestos en la dieta del ganado puede mejorar la salud metabólica de los animales y aumentar la densidad nutricional general de la carne y la leche. Para fundamentar mejor las prácticas de pastoreo, se necesita una investigación más exhaustiva que establezca posibles vínculos entre los indicadores de la salud del suelo, de los animales y de las plantas.
Datos preliminares y objetivos del estudio. Los objetivos de este estudio fueron: 1) determinar las posibles diferencias en los perfiles de metabolitos de la carne de vacuno alimentado con pasto y con pienso; 2) evaluar si existen asociaciones entre estos perfiles y la salud del suelo y la abundancia de nutrientes; y 3) evaluar los biomarcadores de salud en los animales para comprender mejor los posibles efectos de los distintos estados nutricionales sobre su salud y la composición nutricional de la carne [11].
Resultados. Se observaron diferencias notables en la salud del suelo, la composición del pienso y los perfiles metabólicos de la carne de vacuno entre los sistemas de producción basados en pastoreo y los alimentados con cereales. Los suelos de pastoreo mostraron, en general, mayores niveles de materia orgánica, disponibilidad de nutrientes y minerales en comparación con los suelos de los campos de maíz utilizados para la producción de pienso. La metabolómica no dirigida reveló una variación bioquímica sustancial entre el forraje de pastoreo y las raciones mixtas a base de cereales (Figura 20). El pasto contenía niveles más altos de fitoquímicos, fenoles y compuestos relacionados con el omega-3, mientras que las raciones de cereales presentaban niveles más altos de dipéptidos y metabolitos relacionados con el omega-6. Estas diferencias dietéticas se reflejaron en la propia carne. La carne de vacuno alimentado con pasto contenía niveles más altos de compuestos fenólicos, ácidos grasos omega-3, acilcarnitinas de cadena larga, antioxidantes y derivados de la vitamina E y la vitamina A. Por el contrario, la carne de vacuno alimentado con cereales contenía niveles más altos de ciertos dipéptidos, azúcares, vitaminas del grupo B y marcadores de estrés oxidativo (Figura 21).
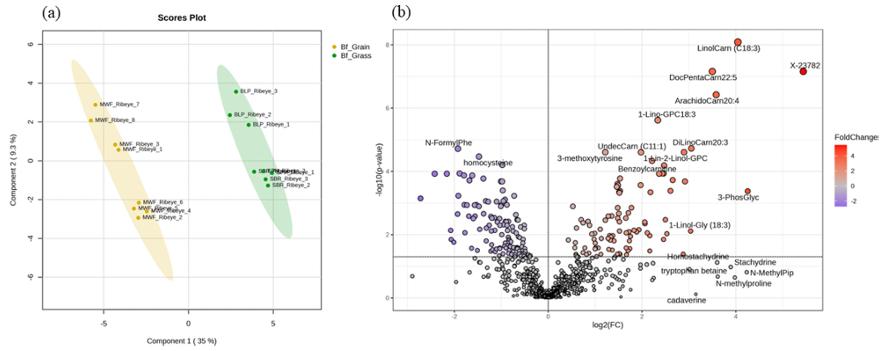
Figura 20. Análisis multivariante y univariante de los perfiles de metabolitos en muestras de carne de vacuno alimentado con pasto y con pienso. (A) Análisis discriminante de mínimos cuadrados parciales (PLS-DA) y (B) gráfico de volcán de los metabolitos diferenciales en muestras de carne de vacuno alimentado con pasto (n = 8) y con pienso (n = 8). Imagen reproducida de Ahsin et al., NPJ Sci Food, 2025, con licencia CC BY 4.0.
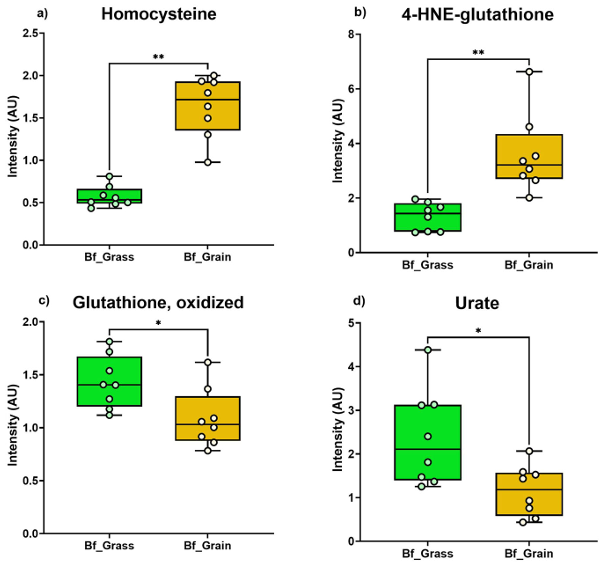
Figura 21. Marcadores de estrés oxidativo y antioxidantes en carne de vacuno alimentado con pasto (Bf_Grass) y con pienso (Bf_Grain). El gráfico de barras muestra la abundancia de marcadores de estrés oxidativo: (A) homocistina, (B) 4-HNE-glutatión, y de antioxidantes: (C) glutatión oxidado, (D) urato, en ambos grupos de ganado. Imagen reproducida de Ahsin et al., NPJ Sci Food, 2025, con licencia CC BY 4.0.
Conclusiones del capítulo
- La metabolómica es una herramienta de gran valor para obtener información biológica que va más allá de lo que pueden aportar otras ciencias «ómicas».
- La información obtenida gracias a la metabolómica puede ser clave para reducir costes y obtener una ventaja competitiva.

Descarga la guía completa en formato PDF
Descargue esta completa guía desarrollada para enseñarle los entresijos de una de las herramientas ómicas más potentes de la caja de herramientas de cualquier científico: la metabolómica.
Descargar ahoraPóngase en contacto con nosotros
Hable con un experto
Solicite un presupuesto para nuestros servicios, obtenga más información sobre tipos de muestras y procedimientos de manipulación, solicite una carta de apoyo o envíe una pregunta sobre cómo la metabolómica puede hacer avanzar su investigación.
Sede social
617 Davis Drive, Suite 100
Morrisville, NC 27560
Dirección postal:
P.O. Box 110407
Research Triangle Park, NC 27709
+1 (919) 572-1711
Sede internacional
Metabolon GmbH
Zeppelinstraße 3
85399 Hallbergmoos
Alemania