Chapitre 3
La métabolomique au service des applications commerciales
Dans ce chapitre, vous découvrirez comment la métabolomique peut apporter des informations fonctionnelles sur les phénotypes pathologiques que les biomarqueurs traditionnels ne permettent pas d'obtenir, comment elle peut identifier des biomarqueurs diagnostiques très performants qui échappent aux autres sciences omiques, et comment elle peut contribuer à l'amélioration de l'alimentation humaine afin de prévenir les maladies et d'améliorer la santé en général.
Vue d'ensemble
La métabolomique est devenue un outil précieux dans divers secteurs d'application, notamment le développement de médicaments, la nutrition humaine, l'élevage et les soins aux animaux de compagnie. Les modifications métaboliques peuvent permettre de mettre en lumière de nouvelles cibles thérapeutiques et de nouveaux mécanismes d'action des médicaments, d'identifier les liens entre l'alimentation et les phénotypes pathologiques, et de révéler des biomarqueurs utiles pour la santé animale. Nous présentons ici quelques études qui montrent comment la métabolomique a contribué à résoudre un problème et/ou a apporté des éclairages scientifiques essentiels sur des thèmes liés à certains secteurs d'application.
Développement de médicaments
Mieux comprendre la toxicité médicamenteuse, là où les biomarqueurs traditionnels ont échoué
Contexte. La dyslipidémie, caractérisée par un taux anormalement élevé de lipoprotéines de basse et de très basse densité (LDL/VLDL) dans le sang, constitue un facteur de risque majeur de maladies cardiovasculaires et d’accidents vasculaires cérébraux. Les agonistes des récepteurs de l'acide nicotinique (NAR) traitent la dyslipidémie en activant les récepteurs des cellules adipeuses afin d'inhiber le métabolisme des triglycérides et de réduire les taux circulants de LDL et de VLDL. Dans le cadre d'une étude, des chercheurs des laboratoires de recherche Merck développaient un agoniste des NAR (SCH 900424) pour traiter la dyslipidémie, mais leurs recherches ont mis en évidence une toxicité potentielle induite par le médicament qu'ils souhaitaient mieux comprendre et surveiller [1].
Données préliminaires et objectifs de l'étude. Lors d'études précliniques, le SCH 900424 a provoqué de manière inattendue une morbidité rapide et une insuffisance rénale aiguë (IRA) chez la souris, ce qui suggère que le médicament agissait à la fois sur le système rénal et sur le système nerveux central (SNC). Les marqueurs traditionnels de la fonction rénale, notamment la créatinine et l'azote uréique sanguin (BUN), n'étaient pas suffisamment sensibles pour prédire ou caractériser la toxicité induite par le médicament, et le ou les mécanismes sous-jacents étaient inconnus. Les objectifs de cette étude étaient d'identifier des biomarqueurs sensibles et précoces des toxicités liées au SCH 900424 chez la souris et d'élucider le ou les mécanismes ayant conduit à cette toxicité.
Méthodes. Des souris CD-1 mâles adultes ont reçu des doses faibles, moyennes et élevées de SCH 900424 ou d’un agoniste témoin. Des échantillons de plasma, d'urine et de cerveau ont été prélevés 1, 4, 8 et 24 heures après l'administration. Les échantillons ont été traités puis analysés à l'aide de la métabolomique globale afin d'identifier les perturbations dans les voies biochimiques pertinentes. La métabolomique ciblée a ensuite été utilisée pour mesurer les composés biochimiques d'intérêt identifiés lors des analyses de découverte globales. Trois matrices d'échantillons ont été analysées afin de garantir une étude mécanistique exhaustive des pathologies liées à l'IRA et au SNC.
Résultats. Dans le plasma, le sulfate de 3-indoxyle (3IS), une toxine urémique d'origine intestinale issue du métabolisme du tryptophane, s'est révélé être l'un des principaux biomarqueurs de l'IRA induite par le SCH 900424. Ce marqueur a augmenté de manière significative chez les souris traitées par rapport aux témoins et a également augmenté plus tôt et plus fortement que la créatinine ou l'urée (Figure 5, dose élevée), ce qui montre que le 3IS est un marqueur de l'IRA plus sensible que les marqueurs traditionnels de la fonction rénale.
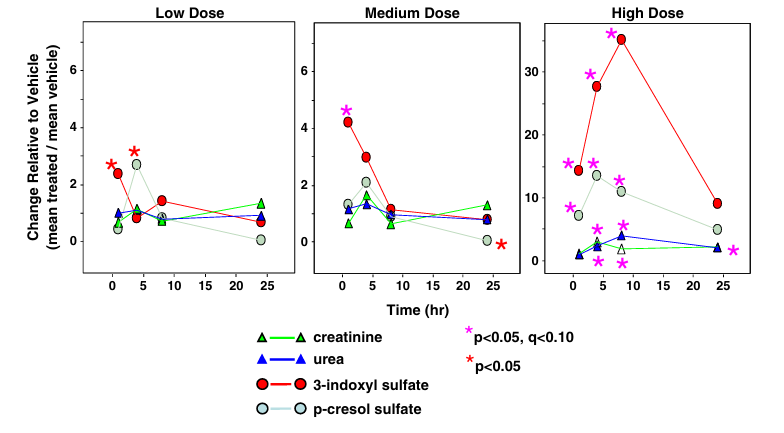
Figure 5. Variations relatives par rapport au véhicule dans le plasma pour le 3IS, le PCS, l'urée et la créatinine après administration du SCH 900424. Sur la base d'une analyse métabolomique globale exhaustive, le 3IS et le sulfate de p-crésol ont été identifiés comme les marqueurs plasmatiques les plus significatifs sur le plan statistique de la toxicité rénale dans cette étude et se sont révélés plus sensibles que l'urée et la créatinine. Image reproduite à partir de Zgoda-Pols et al., Toxicol Appl Pharmacol, 2011, sous licence CC BY 4.0.
Les taux plasmatiques de sulfate de p-crésol, une toxine urémique issue du métabolisme de la tyrosine et de la phénylalanine, étaient également significativement élevés (Figure 5, dose élevée). Plusieurs acides aminés, dont le tryptophane, ont présenté une diminution significative dans le plasma, tandis que de nombreux catabolites du tryptophane, notamment la kynurénine, le kynurénate et l'indole-3-lactate, ont affiché une augmentation significative (Figure 6). Dans l'urine, des augmentations significatives du glucose et des acides aminés ont été observées. Dans des conditions normales, la quasi-totalité des acides aminés et des sucres filtrés sont réabsorbés dans les tubules proximaux. Cependant, une lésion rénale altérerait la réabsorption et entraînerait leur excrétion dans l'urine. La signature métabolique observée dans le plasma et l'urine confirmait un véritable dysfonctionnement rénal plutôt que des modifications métaboliques isolées.
Une autre conclusion importante de l'étude a été l'élévation significative et dose-dépendante du taux de 3IS dans le tissu cérébral après l'administration du SCH 900424. Cette augmentation s'est accompagnée d'une réduction significative des taux urinaires de 3IS, ce qui indique une altération de la clairance entraînant son accumulation dans le cerveau. Dans l'ensemble, ces résultats suggèrent que l'accumulation de 3IS dans le plasma et le cerveau pourrait non seulement contribuer à l'insuffisance rénale aiguë, mais aussi à une toxicité du SNC et à une morbidité rapide chez la souris.
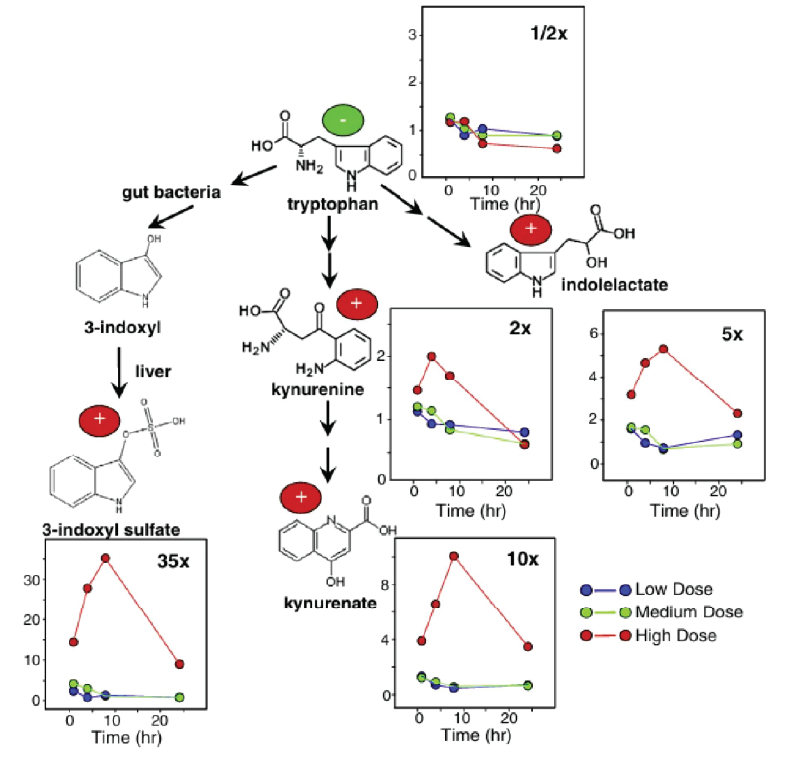
Figure 6. Voies cataboliques du tryptophane avec les variations relatives par rapport au véhicule pour chaque catabolite. Alors que le tryptophane diminuait au fil du temps avec les doses les plus élevées de SCH 900424, ses catabolites, et en particulier le 3IS, ont augmenté de manière spectaculaire. Image reproduite à partir de Zgoda-Pols et al., Toxicol Appl Pharmacol, 2011, sous licence CC BY 4.0.
Conclusions de l'étude
- Les résultats de cette étude suggèrent que le 3IS constitue un marqueur précoce et plus sensible de la toxicité rénale induite par le SCH 900424 que les marqueurs chimiques traditionnels. Dans la pratique, cela pourrait améliorer la détection précoce de la néphrotoxicité lors du développement de médicaments.
- L'accumulation observée de toxines urémiques d'origine intestinale, la perturbation du métabolisme du tryptophane et la diminution de l'excrétion urinaire ont fourni des preuves mécanistiques d'une accumulation de toxines urémiques due à un dysfonctionnement rénal.
- L'accumulation de 3IS dans le tissu cérébral, et non pas uniquement dans le plasma, suggère l'existence d'un lien mécanistique entre l'insuffisance rénale et la survenue rapide de complications chez la souris ; il s'agit là d'une nouvelle hypothèse qui pourra être vérifiée dans le cadre d'études futures.
- Cette étude montre comment la métabolomique peut contribuer à transformer une observation phénotypique non caractérisée en un profil de biomarqueurs fondé sur les mécanismes sous-jacents, afin d'orienter la sélection de candidats-médicaments avec plus de rapidité et de précision.
Sauver un programme de développement de médicaments
Contexte. L'infection à C. difficile (ICD) provoque une colite inflammatoire suivie d'une diarrhée très invalidante pour les patients. Même sous traitement antibiotique, 20 à 30 % des patients présentent des ICD récurrentes en raison d'une perturbation persistante de leur microbiome. Dans un intestin sain, les bactéries intestinales inhibent la croissance et la germination de C. difficile en transformant les acides biliaires primaires conjugués en acides biliaires secondaires et en produisant des acides gras à chaîne courte et moyenne. Il a été démontré que les spores du microbiote fécal produites par des membres du phylum Firmicutes recolonisent l'intestin avec des bactéries qui protègent contre les CDI récurrentes. Dans une étude, des chercheurs de Seres Therapeutics ont mis au point le VOS (anciennement SER-109), un produit biothérapeutique vivant à base de microbiome, administré par voie orale, destiné au traitement et à la prévention des CDI récurrentes [2].
Données préliminaires et objectifs de l'étude. La prise de VOS s'est avérée régulière chez les patients et seuls 13,3 % d'entre eux ont présenté une récidive d'infection à Clostridioides difficile (ICD) sur l'ensemble des doses administrées dans le cadre d'une étude de phase I en ouvert. Cependant, le VOS n'a pas atteint son critère d'efficacité principal lors d'une étude de phase II randomisée et contrôlée par placebo menée en suivi, dans laquelle il a été administré à une dose fixe ayant démontré son efficacité en phase I. En réponse, ce groupe a mené plusieurs études métabolomiques de suivi. L'objectif de leur première étude de suivi était de concilier ces résultats disparates en évaluant les modifications induites par le VOS sur le microbiome et les métabolites associés aux microbes [3]. Leur deuxième objectif est décrit dans la section suivante intitulée « Succès dans un essai de phase 3 malgré l'échec en phase 2 ».
Méthodes. Dans les essais cliniques de phase 1 et 2, la prise de VOS a été évaluée en comparant le nombre d'espèces bactériennes associées au VOS dans des échantillons de selles prélevés 1, 4 et 8 semaines après l'administration, et la récidive de l'infection à C. difficile a été déterminée sur la base des symptômes cliniques. Un séquençage métagénomique complet de type « shotgun » et des analyses métabolomiques globales ont été réalisés a posteriori sur les ensembles de données des phases 1 et 2 afin d'évaluer l'impact du VOS sur les résultats cliniques.
Résultats. Dans l'essai clinique de phase 2, les participants traités qui n'ont pas développé de récidive de CDI présentaient un nombre significativement plus élevé d'espèces de VOS une semaine après l'administration du traitement que ceux ayant connu une récidive. Une analyse post hoc des résultats des essais de phase 1 et de phase 2 a été réalisée afin d'évaluer l'ampleur de la prise de greffe une semaine après l'administration d'une dose élevée (phase 1), faible (phase 1) ou fixe (phase 2) aux participants. Les sujets de la phase 1 ayant reçu la dose élevée présentaient un nombre significativement plus élevé d'espèces de VOS à la semaine 1 que les deux autres cohortes de dosage (Figure 7). De plus, la prise de greffe à la semaine 1 s'est avérée très variable chez les participants de la phase 2, ce qui suggère que la réussite de la prise de greffe dépendait de la dose et que le dosage était sous-optimal dans l'essai de phase 2.
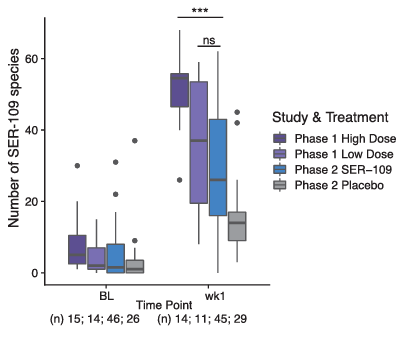
Figure 7. Relation entre la colonisation par des espèces de VOS et les doses administrées dans les études de phase 1 et 2. Les participants ayant reçu la dose élevée dans l'étude de phase 1 présentaient un nombre significativement plus élevé d'espèces de VOS que les participants du groupe de traitement de l'étude de phase 2 ayant reçu une dose fixe faible (p < 0,001). Image reproduite à partir de McGovern et al., Clin Infect Dis, 2021, sous licence CC BY 4.0.
Il est intéressant de noter qu'il existait également une corrélation positive significative entre le nombre d'espèces de VOS et l'abondance des acides biliaires secondaires, à savoir l'acide lithocholique (LCA) et l'acide désoxycholique (DCA), ce qui suggère qu'une implantation accrue de ces espèces entraîne une conversion accrue des acides biliaires primaires conjugués en acides biliaires secondaires, une étape clé pour freiner la croissance de C. difficile et rétablir une santé intestinale normale (figure 8).
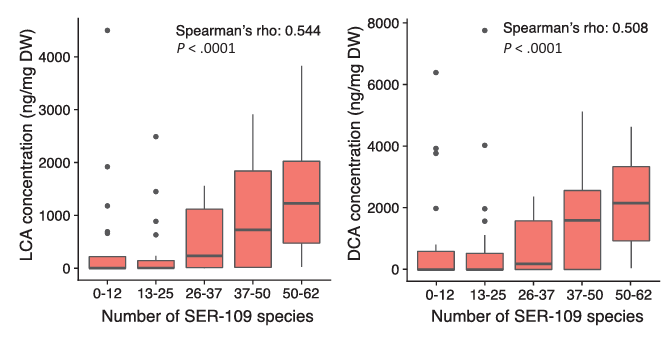
Figure 8. Relation entre la prise de greffe des souches VOS (anciennement SER-109) et la concentration en acides biliaires secondaires dans l'étude de phase 2. Le nombre d'espèces de VOS une semaine après l'administration était significativement associé à la concentration de LCA et de DCA. Abréviations : DCA - acide désoxycholique, DW - poids sec, LCA - acide lithocholique. Image reproduite à partir de McGovern et al., Clin Infect Dis, 2021, sous licence CC BY 4.0.
Conclusions de l'étude
- Cette étude a montré que l'échec de l'essai clinique de phase 2 était probablement dû à un dosage sous-optimal, qui a entraîné une prise de greffe inégale, plutôt qu'à une faible efficacité thérapeutique.
- Ces résultats ont également conduit à émettre l'hypothèse selon laquelle la colonisation par VOS entraîne des changements fonctionnels importants par l'intermédiaire de métabolites clés associés aux microbes, qui brisent le cycle des infections récurrentes à C. difficile.
- Dans cette première étude de suivi, la métabolomique a joué un rôle déterminant pour expliquer pourquoi les résultats des études de phase 1 et de phase 2 divergeaient, ce qui a permis à l'équipe de poursuivre un essai de phase 3 repensé malgré l'échec de la phase 2, évitant ainsi qu'un traitement prometteur ne soit abandonné et préservant l'investissement commercial. L'étude de cas suivante présente les résultats de l'essai de phase 3 mené par ce groupe.
Un essai de phase III couronné de succès malgré l'échec de la phase II
Données préliminaires et objectifs de l'étude. À la lumière des résultats décrits dans l'étude de cas ci-dessus, les chercheurs ont repensé l'étude de phase III afin d'adopter une stratégie posologique plus efficace [4]. Les participants du groupe de traitement ont reçu une dose fixe de VOS dix fois supérieure à celle administrée lors de la phase II, dans le but d'évaluer l'efficacité thérapeutique du VOS dans des conditions propices à la prise de greffe.
Méthodes. L'efficacité a été évaluée sur la base de la réduction du risque relatif et de la réponse clinique durable jusqu'à 8 semaines après l'administration du traitement. Plus précisément, le risque relatif a été défini comme le pourcentage de patients présentant une récidive dans le groupe VOS divisé par le pourcentage correspondant dans le groupe placebo. La réponse clinique durable a été définie comme l'absence de récidive pendant les 8 semaines de suivi des patients. Des analyses métabolomiques globales ont été réalisées sur des échantillons de selles prélevés 1, 2 et 8 semaines après l'administration du traitement afin d'évaluer les profils d'acides biliaires dans l'intestin.
Résultats. Il a été démontré que le VOS (anciennement SER-109) était plus efficace que le placebo pour réduire le risque de récidive de l'infection à C. difficile et pour induire une réponse clinique durable chez les patients (figure 9).
Le groupe traité par VOS a présenté une augmentation significativement plus importante des acides biliaires secondaires par rapport aux valeurs initiales, comparé au groupe placebo (figure 10), ce qui concorde avec la diminution des quantités de bactéries entérobactéries pro-inflammatoires et l'augmentation des bactéries Firmicutes, espèces considérées comme des régulateurs clés de la CDI.
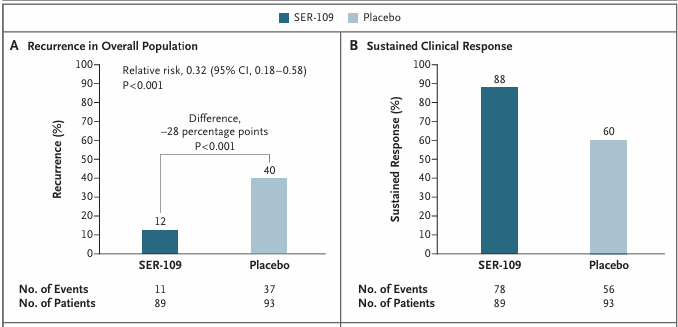
Figure 9. Récidive de l'infection à C. difficile jusqu'à 8 semaines. (A) L'analyse d'efficacité principale a montré la supériorité du VOS (SER-109) par rapport au placebo en termes de réduction du risque de récidive de l'infection à C. difficile. (B) Le VOS s'est également révélé supérieur au placebo pour induire une réponse clinique durable. Image reproduite à partir de Feuerstadt et al., N Engl J Med, 2022, sous licence CC BY 4.0.
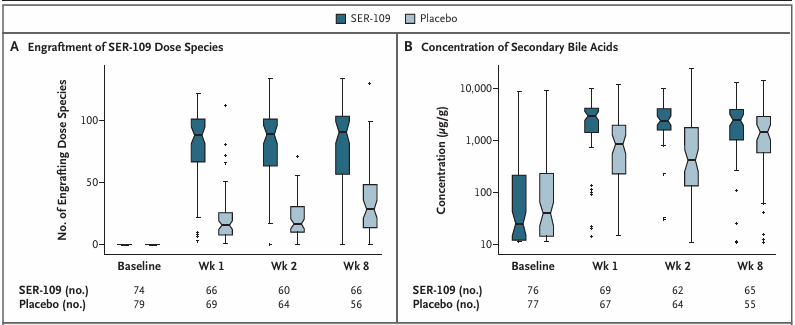
Figure 10. Modifications de la composition et du profil métabolomique du microbiome après administration de VOS (SER-109) ou d’un placebo. (A) Nombre d’espèces ayant pris pied et (B) concentration des acides biliaires secondaires groupés détectés dans les échantillons de selles des patients 1, 2 et 8 semaines après l’administration de VOS ou d’un placebo. Image reproduite à partir de Feuerstadt et al., N Engl J Med, 2022, sous licence CC BY 4.0.
Conclusions de l'étude
- Un essai de phase III repensé pour optimiser la posologie sur la base de données métabolomiques a permis au VOS de s'implanter correctement et, par conséquent, de démontrer sa supériorité par rapport au placebo dans la prévention de la récidive de l'infection à Clostridioides difficile.
- Les données métabolomiques issues de cette étude et des études cliniques précédentes suggèrent que la prise de greffe du VOS dépend de la dose, et que son mécanisme d'action pourrait être lié à la production d'acides biliaires secondaires.
- Ces résultats ont conduit à une hypothèse tout à fait plausible concernant le mécanisme d'action du VOS. Étant donné qu'il est essentiel de comprendre le mécanisme d'action d'un agent thérapeutique pour optimiser l'efficacité du traitement, anticiper les effets indésirables et déterminer les posologies appropriées, cette étude pourrait faciliter la prise de décisions rentables pour les études cliniques de suivi
À la découverte de biomarqueurs diagnostiques hautement performants qui échappent aux autres sciences omiques
Introduction. Les taux de survie au cancer colorectal (CCR) pourraient être améliorés grâce à un dépistage précoce ; il existe donc un besoin important de biomarqueurs non invasifs pour faciliter ce dépistage. Les modifications du métabolisme énergétique sont une caractéristique de la tumorigenèse, car les cellules cancéreuses dépendent de la glycolyse aérobie pour produire de l'énergie et des intermédiaires glycolytiques, qui servent de précurseurs aux lipides et aux acides aminés. Les différences dans les profils plasmatiques et sériques des acides aminés et des lipides ont démontré une sensibilité et une spécificité élevées pour distinguer les patients atteints de CCR des témoins sains.
Données préliminaires et objectifs de l'étude. De plus en plus de données suggèrent que les plaquettes contribuent à la progression du cancer. Des expériences de co-culture ont montré que le plasma riche en plaquettes (PRP) favorise un phénotype prolifératif chez les cellules cancéreuses, et on pense que l'adhésion des plaquettes facilite les métastases tumorales. Dans cette étude, les chercheurs ont émis l'hypothèse que le PRP présentant une activité biologique élevée pourrait révéler des changements métaboliques susceptibles d'aider au dépistage précoce du CCR. L'objectif de cette étude était d'analyser les profils métaboliques du plasma riche en plaquettes (PRP) chez des patients atteints de CCR et des témoins sains afin d'identifier les changements métaboliques pouvant aider à détecter le CCR à un stade précoce [5].
Méthodes. Une analyse métabolomique globale a été réalisée sur du PRP prélevé et regroupé selon les catégories suivantes : patients atteints d'un CCR de stade I et II, patients atteints d'un CCR de stade III et IV, et témoins sains. Une analyse par grappes et une analyse d'enrichissement par classe ont été utilisées pour évaluer les différences de profils entre les trois cohortes.
Résultats. Les taux de lactose, de glycérol-3-phosphate, d’acide 2-hydroxyglutarique, d’acide isocitrique et d’acide citrique étaient significativement plus élevés dans le PRP prélevé chez les patients atteints d’un CCR par rapport aux témoins (figure 11). Bien que les taux de ces métabolites ne différaient pas de manière significative entre les tumeurs malignes à un stade précoce (stades I et II) et celles à un stade avancé (stades III et IV), leurs concentrations systématiquement plus élevées par rapport aux témoins ne les excluaient pas comme biomarqueurs candidats pour la détection précoce du CCR.
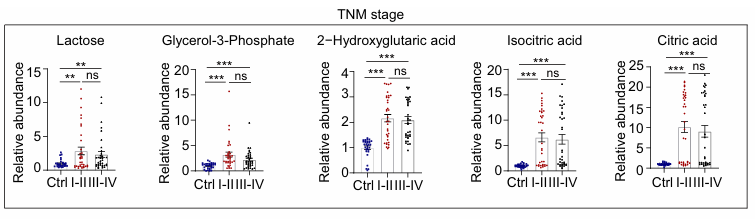
Figure 11. Abondance relative de cinq métabolites glucidiques clés aux différents stades de la progression du CCR. Ces métabolites présentaient des différences significatives entre les patients atteints de CCR et le groupe témoin. Aucune différence n'a été observée entre les tumeurs malignes à un stade précoce (stades I et II) et celles à un stade avancé (stades III et IV). Image reproduite d'après Hu et al., Mol Oncol, 2025, sous licence CC BY 4.0.
Les performances diagnostiques de cette signature composée de 5 métabolites ont été comparées à celles de l'antigène carcino-embryonnaire (CEA) et de l'antigène carbohydrate 19-9 (CA 19-9), deux biomarqueurs sanguins classiques du CCR. Le CEA a présenté une sensibilité de 80 %, une spécificité de 76,67 % et une valeur de l'aire sous la courbe ROC (AUC) de 0,783 ; le CA 19-9 a atteint une sensibilité de 74 %, une spécificité de 63 % et une AUC de 0,641. En revanche, la signature métabolomique a obtenu de bien meilleurs résultats, distinguant le CCR des témoins sains avec une sensibilité de 90 %, une spécificité de 96,67 % et une AUC de 0,961. (Figure 12).
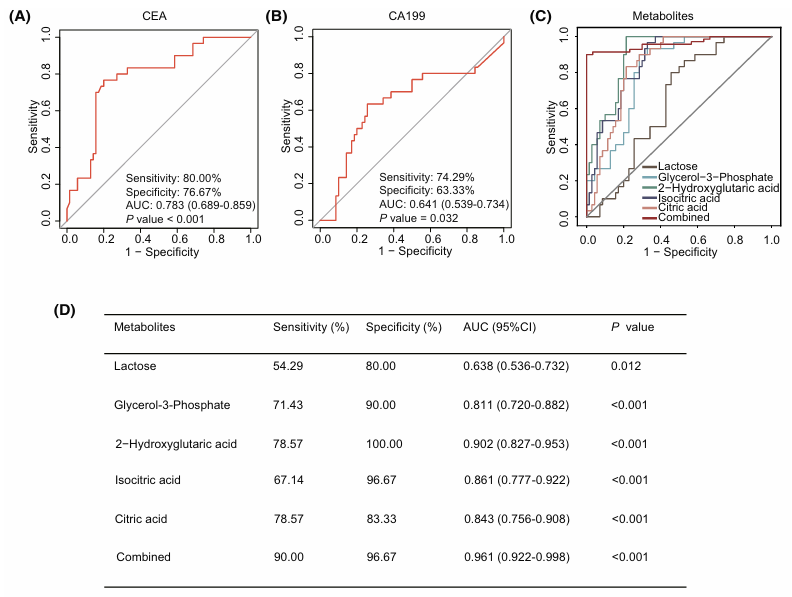
Figure 12. Performances diagnostiques de la signature métabolique par rapport aux biomarqueurs traditionnels du CCR. (A) Efficacité diagnostique du CEA, (B) du CA 19-9, et (C) de chaque métabolite de la signature, pris isolément et en combinaison. (D) Sensibilité, spécificité et aire sous la courbe (AUC) de chaque métabolite de la signature pour distinguer les patients atteints de CCR des témoins sains. Image reproduite à partir de Hu et al., Mol Oncol, 2025, sous licence CC BY 4.0.
Nutrition humaine
Identification des liens mécanistiques entre l'alimentation et les maladies rénales chroniques
Introduction. L'insuffisance rénale chronique (IRC) touche 800 millions de personnes dans le monde et engendre un coût annuel d'environ 407 milliards de dollars. L'alimentation a un impact significatif sur la santé rénale et, d'après des résultats récents, les régimes alimentaires à base de végétaux pourraient constituer un moyen rentable de prévenir l'IRC. L'étude Adventist Health Study-2 (AHS-2) est une étude pluriannuelle visant à évaluer les liens entre le mode de vie, l'alimentation et les maladies. Des résultats récents issus de cette cohorte ont montré un lien entre les régimes alimentaires végétariens et une incidence plus faible du diabète et de l'hypertension, deux facteurs de risque majeurs de l'IRC. Le régime végétarien était également fortement associé à une réduction de la mortalité due aux maladies rénales. Malgré ces liens étroits, on ne savait pas exactement comment les régimes alimentaires végétariens semblaient protéger la santé rénale.
Données préliminaires et objectifs de l'étude. Des analyses métabolomiques antérieures menées au sein de la cohorte ASH-2 ont montré que les végétaliens présentaient des taux plasmatiques plus élevés de métabolites dérivés des polyphénols, d'origine microbienne et liés aux vitamines, tandis que les non-végétariens affichaient des taux plus élevés de lipides et d'acides aminés pro-inflammatoires. Dans le but de mieux comprendre les relations de cause à effet potentielles entre le risque de MRC, sa progression et l'alimentation, cette étude a examiné les associations entre les habitudes alimentaires végétariennes, la fonction rénale et les métabolites liés à l'alimentation en utilisant le débit de filtration glomérulaire estimé (DFGe) comme mesure de la santé rénale [6].
Méthodes. 899 participants de la cohorte AHS-2 ont été classés selon leurs habitudes alimentaires, à savoir végétaliens, pesco-végétariens et non végétariens. Une analyse métabolomique globale a été réalisée sur des échantillons de sérum prélevés chez ces participants. Les taux sériques de créatinine ont été utilisés pour calculer le DFG estimé. Pour l'analyse des sous-classes de métabolites, les valeurs individuelles des métabolites composant une sous-classe ont été moyennées afin d'obtenir une moyenne composite pour cette sous-classe. Les scores T ont été obtenus en divisant l'écart-type des moyennes composites, en tenant compte des covariances entre les métabolites. Le nombre de métabolites significatifs ou différentiels pour chaque sous-classe a été déterminé, ainsi que le nombre de métabolites différentiels associés de manière positive ou inverse au régime alimentaire.
Résultats. De nombreux métabolites ont été associés de manière spécifique au régime alimentaire et aux taux de créatinine, ce qui a mis en évidence des liens précis entre les habitudes alimentaires et le phénotype métabolique. Par exemple, par rapport aux non-végétariens, les végétaliens présentaient des taux significativement plus faibles de certaines toxines urémiques, notamment le p-crésol, le sulfate de 3-méthylcatéchol et le glucuronide, qui sont associés à un eGFR plus faible et, par conséquent, à une fonction rénale réduite. Des analyses plus approfondies ont permis d'identifier des sous-classes de métabolites (plutôt que des métabolites individuels) associées à différents régimes alimentaires et à la créatinine sérique. Les comparaisons entre les régimes alimentaires végétaliens et non végétariens ont montré que les plasmalogènes et les lysoplasmalogènes représentaient la plus grande proportion de métabolites significatifs dans l'un ou les deux groupes (Figure 13). Il a été démontré que les plasmalogènes et les lysoplasmalogènes jouent un rôle dans la réparation membranaire lors du stress oxydatif. Une diminution des taux circulants de ces sous-classes de métabolites a été observée chez les patients atteints d'IRC avancée, probablement en raison d'un stress oxydatif élevé.
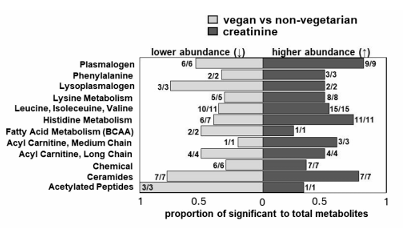
Figure 13. Sous-classes différentielles présentant des associations opposées entre les régimes alimentaires, la créatinine sérique et les métabolites plasmatiques. Les sous-classes présentant une abondance différentielle ont été déterminées après calcul de la moyenne composite (moyenne des métabolites constitutifs) pour chaque groupe alimentaire, puis des scores t. Les barres reflètent la proportion de métabolites différentiels par rapport au nombre total de métabolites du panel associés positivement à la créatinine (à droite de 0) ou inversement au régime alimentaire indiqué (à gauche de 0). Les ratios représentent le nombre de métabolites significatifs associés dans la même direction (inversement ou positivement) par rapport au nombre total de métabolites significatifs dans la sous-classe. La direction prédominante de l'évolution de l'abondance pour chaque sous-classe, reflétant la direction générale des métabolites composants, est représentée graphiquement pour les comparaisons de la créatinine entre le régime végétalien et le régime non végétarien. Image reproduite à partir de Butler et al., J Ren Nutr, 2025, sous licence CC BY 4.0.
Conclusions de l'étude
- Cette étude a mis en évidence des modifications métaboliques propres à différents régimes alimentaires, afin de mettre en lumière les liens mécanistiques entre l'alimentation et le DFG estimé.
- La métabolomique a permis aux chercheurs d'aller au-delà de la simple mesure de la créatinine pour mettre en évidence les changements métaboliques sous-jacents, apportant ainsi des preuves plus solides du lien direct entre les voies métaboliques influencées par l'alimentation et la santé rénale.
Surmonter les limites des auto-évaluations alimentaires grâce à la normalisation des empreintes biomarqueurs de la consommation alimentaire
Introduction. Les méthodes traditionnelles d'évaluation de l'alimentation, qui reposent souvent sur les déclarations des participants, sont sujettes à des erreurs et à des biais. La métabolomique permet de pallier cette limite en identifiant les composés d'origine alimentaire afin de fournir une empreinte moléculaire objective de l'apport alimentaire. Bien que de nombreux biomarqueurs alimentaires aient été identifiés, la plupart des études se sont concentrées sur des aliments individuels plutôt que sur des régimes alimentaires globaux, et rares sont les essais alimentaires contrôlés ayant adopté une approche croisée randomisée pour réduire l'hétérogénéité lors de la quantification des associations avec des régimes alimentaires spécifiques.
Données préliminaires et objectifs de l'étude. Afin de combler les lacunes existant dans les rapports métabolomiques alimentaires actuels, cette étude a mené un essai alimentaire randomisé, croisé et contrôlé comparant un régime alimentaire australien sain (HAD), fondé sur les recommandations alimentaires australiennes en vigueur, à un régime alimentaire australien typique (TAD). Les objectifs des chercheurs étaient 1) de décrire les changements métaboliques dans le plasma et l'urine en réponse à ces deux régimes alimentaires, et 2) d'identifier les métabolites discriminants spécifiques au régime alimentaire sain afin de développer un score de qualité alimentaire dérivé du métabolome qui pourrait aider à évaluer l'association entre le régime alimentaire sain et les marqueurs de la santé cardiométabolique [7].
Méthodes. Dans le cadre d'un essai alimentaire croisé randomisé de 8 semaines, des adultes en bonne santé ont suivi deux interventions alimentaires contrôlées de 2 semaines chacune, l'une consistant en un régime alimentaire sain et l'autre en un régime alimentaire habituel, séparées par une période de sevrage. Tous les repas ont été fournis afin de garantir l'observance et de minimiser la variabilité alimentaire. Des échantillons sanguins et urinaires à jeun ont été prélevés avant et après chaque intervention, puis analysés à l'aide d'un profilage métabolomique non ciblé. Une régression élastique a été utilisée pour identifier les métabolites permettant de distinguer le mieux le régime alimentaire sain du régime alimentaire habituel. Ces métabolites ont servi à établir un score composite de qualité alimentaire dérivé du métabolome, qui a ensuite été testé pour déterminer ses associations avec des marqueurs cardiométaboliques, notamment la pression artérielle, les lipides et la glycémie à jeun.
Résultats. Au bout de deux semaines, 408 métabolites plasmatiques et 319 métabolites urinaires présentaient des différences par rapport aux valeurs de référence, dont un grand nombre appartenaient aux voies métaboliques des lipides, des acides aminés et des xénobiotiques. La comparaison directe des deux régimes alimentaires a permis d’identifier des centaines de métabolites présentant des différences significatives. Parmi ceux-ci, 43 ont montré des variations cohérentes tant dans le plasma que dans l’urine, ce qui suggère que chaque régime alimentaire a généré une signature métabolique bien définie (figure 14). Une analyse par apprentissage automatique a permis d'identifier 31 métabolites plasmatiques et 34 métabolites urinaires qui distinguaient le mieux le régime alimentaire sain (HAD) du régime alimentaire typique (TAD). Ces métabolites ont été utilisés pour créer un score de qualité alimentaire dérivé du métabolome (Figure 15 A, B). Des scores plus élevés, indiquant une adhésion étroite au régime HAD, étaient significativement associés à une amélioration des marqueurs cardiométaboliques, notamment une baisse de la pression artérielle, des triglycérides, du cholestérol LDL et de la glycémie à jeun (Figure 15 C-H).
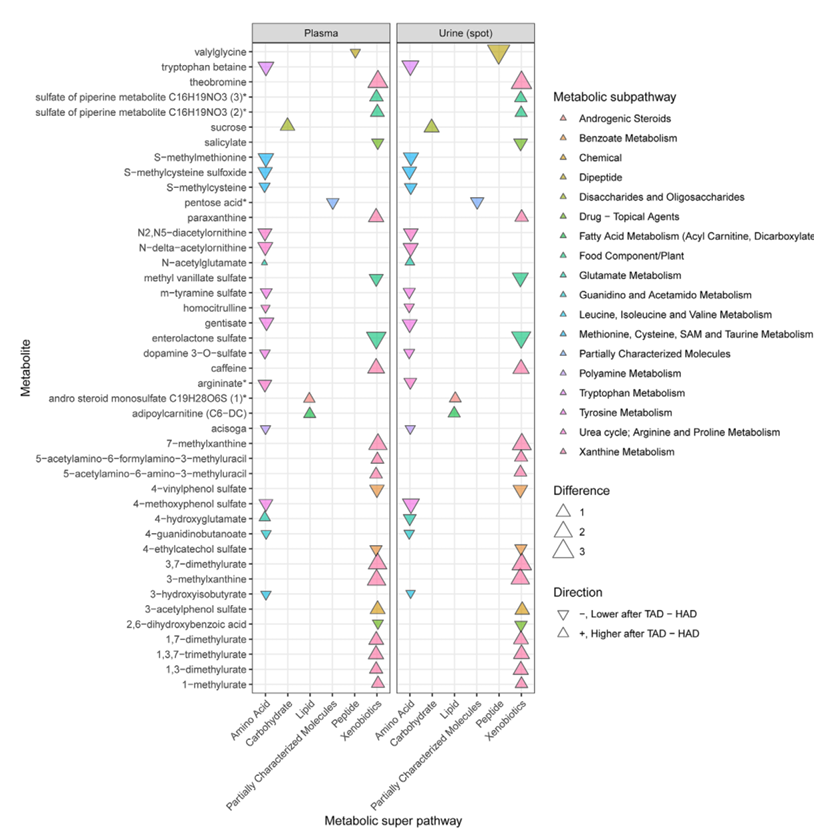
Figure 14. Amplitude et sens de la variation de 43 métabolites après deux semaines de consommation d'un régime HAD par rapport à un régime TAD. Les concentrations de ces métabolites présentaient des différences systématiques entre les groupes, tant dans le plasma que dans l'urine. Image reproduite à partir de Stanford et al., Mol Nutr Food Res, 2025, sous licence CC BY 4.0.
Figure 15. Élaboration d’un score biomarqueur de la qualité de l’alimentation dérivé du métabolome et son association avec des marqueurs de la santé cardiométabolique. Les métabolites discriminants et leurs coefficients issus (A) du plasma et (B) de l’urine sont présentés. Ces coefficients ont été utilisés pour établir un score composite de qualité alimentaire, où un score plus élevé était associé à une amélioration significative des paramètres cardiométaboliques, notamment (C) une pression artérielle systolique plus basse, (D) une pression artérielle diastolique plus basse, (E) des triglycérides plus bas, (F) un taux de cholestérol LDL plus bas, (G) un taux de cholestérol HDL plus bas et (H) une glycémie à jeun plus basse. Image reproduite à partir de Stanford et al., Mol Nutr Food Res, 2025, sous licence CC BY 4.0.
Conclusions de l'étude
- Dans cette étude, les chercheurs ont eu recours à la métabolomique pour établir un score permettant de mesurer quantitativement l'adhésion au régime alimentaire, ce qui permet de pallier les biais et les erreurs inhérents à l'auto-évaluation de l'apport alimentaire. À l'avenir, ce score de qualité alimentaire pourrait améliorer considérablement l'interprétabilité et la reproductibilité des études nutritionnelles, et par conséquent, optimiser l'efficacité et la rentabilité des futurs protocoles de recherche.
Développement d'outils visant à améliorer la mesure de la consommation d'aliments ultra-transformés
Introduction. Les aliments ultra-transformés (AUT) se caractérisent par la présence d’ingrédients industriels et d’additifs qui ne sont généralement pas utilisés dans la cuisine familiale, notamment des édulcorants artificiels et des amidons modifiés. Bien que la transformation alimentaire joue un rôle important dans la sécurité alimentaire et la durée de conservation des produits, des études observationnelles ont systématiquement établi un lien entre une consommation élevée d’AUT et un risque accru d’obésité, de maladies cardiovasculaires, de cancer et de mortalité toutes causes confondues. À l'heure actuelle, les études nutritionnelles portant sur les UPF sont limitées par un manque d'outils permettant de mesurer avec précision la consommation de ces produits. Les questionnaires courants d'évaluation alimentaire et les rappels alimentaires sur 24 heures ne fournissent pas les informations détaillées sur les ingrédients et la transformation nécessaires pour classer les aliments selon les systèmes de classification de l'industrie.
Données préliminaires et objectifs de l'étude. L'objectif de cette étude était de répondre au besoin de biomarqueurs objectifs permettant de mesurer de manière fiable la consommation d'aliments ultra-transformés (AUT) et ses effets métaboliques, dans le but à long terme d'améliorer l'étude des relations entre l'alimentation et les maladies [8].
Méthodes. Les essais alimentaires contrôlés, dans lesquels tous les aliments sont fournis et la consommation est étroitement surveillée, constituent le cadre idéal pour identifier les biomarqueurs pertinents des aliments ultra-transformés (AUT). La présente étude est une analyse secondaire d’un essai alimentaire contrôlé, randomisé et croisé, au cours duquel 20 adultes ont suivi soit un régime riche en AUT, soit un régime totalement exempt d’AUT. Les régimes étaient appariés en termes de calories et de nutriments. Des échantillons de sang et d'urine ont été prélevés à plusieurs moments tout au long de l'étude, puis analysés par profilage métabolomique global. Les niveaux de métabolites ont été comparés entre les régimes à l'aide de modèles linéaires à effets mixtes, et des analyses supplémentaires ont évalué les changements au niveau des voies métaboliques.
Résultats. Par rapport au régime témoin, le régime riche en aliments ultra-transformés (UPF) a induit des modifications métaboliques généralisées et systématiques. Comme prévu, plusieurs métabolites directement liés aux additifs alimentaires et aux conservateurs ont connu une augmentation significative dans le régime riche en UPF, tandis que les métabolites associés aux fruits et légumes entiers ont diminué, reflétant une consommation moindre de ces aliments. Des changements ont également été observés dans les voies métaboliques liées au métabolisme des lipides, aux acides biliaires, aux acides aminés et aux métabolites du microbiote intestinal, montrant que la consommation d'UPF modifie à la fois le métabolisme de l'hôte et celui lié au microbiote. Il est important de noter que bon nombre de ces changements métaboliques étaient cohérents entre le plasma et l'urine, ce qui confirme leur potentiel en tant que biomarqueurs fiables de la consommation d'UPF et de la réponse métabolique à court terme (Figure 16).
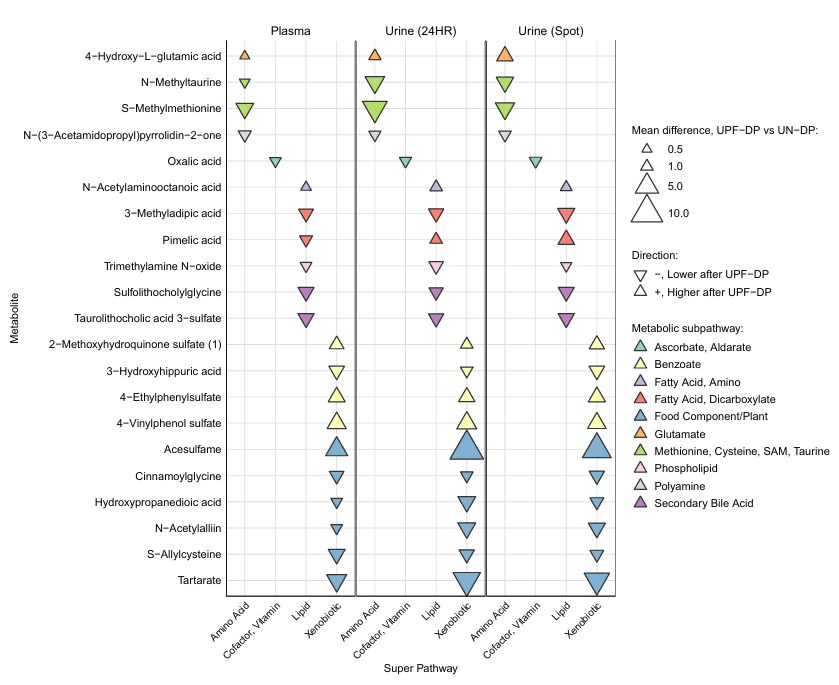
Figure 16. Amplitude et sens des différences moyennes observées à la deuxième semaine au niveau des métabolites après la consommation d’un régime riche en aliments ultra-transformés (UPF) par rapport à un régime témoin. Les métabolites répertoriés ici (n = 21) présentaient des différences chez les participants individuels (n = 20) entre les groupes UPF-DP et UN-DP à la deuxième semaine pour le plasma, ainsi qu’à la première et à la deuxième semaine pour l’urine de 24 heures. Les différences moyennes entre les métabolites ont été estimées à l'aide d'un modèle linéaire mixte ajusté en fonction du régime alimentaire, de la phase, de la séquence et du moment de prélèvement pour l'urine uniquement, ainsi que de l'apport énergétique (EI) calculé au cours de la semaine précédant le prélèvement, avec des interceptions aléatoires spécifiques à chaque sujet. Image reproduite à partir de O'Connor et al., J Nutr, 2023, sous licence CC BY 4.0.
Conclusions de l'étude
- Cette étude montre qu'une alimentation riche en aliments ultra-transformés entraîne des modifications nettes et mesurables du métabolome plasmatique et urinaire. Les métabolites directement liés aux aliments ultra-transformés, notamment les conservateurs à base de benzoate et les édulcorants artificiels, ainsi que ceux associés à des modifications du microbiome, pourraient servir de biomarqueurs objectifs de la consommation d'aliments ultra-transformés.
- Ces biomarqueurs pourraient contribuer à pallier certaines limites de longue date des méthodes traditionnelles d'évaluation de l'alimentation et à améliorer la précision et la reproductibilité des études visant à caractériser la relation entre l'alimentation et la santé.
Santé animale et élevage
Caractérisation du mécanisme d'action d'un traitement largement utilisé mais mal compris contre la diarrhée chronique du gros intestin chez le chien
Introduction. La diarrhée chronique du gros intestin chez le chien est un trouble gastro-intestinal courant résultant d’interactions complexes entre le microbiome intestinal, l’alimentation, le métabolisme de l’hôte, les réponses immunitaires et les facteurs environnementaux. Des données récentes suggèrent que la dysbiose microbienne contribue à cette affection en modifiant les produits métaboliques intestinaux, notamment la production de métabolites inflammatoires ou nocifs. L'ajout de fibres à l'alimentation peut améliorer la santé gastro-intestinale en favorisant une fermentation microbienne bénéfique et en augmentant la production de postbiotiques protecteurs, notamment les acides gras à chaîne courte (AGCC), les acides biliaires secondaires et les indoles. Bien que l'intervention nutritionnelle soit considérée comme le traitement de première intention de la diarrhée chronique du gros intestin chez le chien, les mécanismes à l'origine de son effet thérapeutique sont mal compris.
Données préliminaires et objectifs de l'étude. Des chercheurs de Hill’s Pet Nutrition, Inc. ont démontré qu'un mélange exclusif de fibres solubles et insolubles, sélectionnées pour leur activité prébiotique et postbiotique, améliorait les scores relatifs aux selles. L'objectif de cette étude de suivi était d'évaluer l'impact mécanistique d'une intervention alimentaire enrichie en fibres sur la santé gastro-intestinale chez les chiens souffrant activement de diarrhée du gros intestin [9].
Méthodes. Les chercheurs ont mené un essai clinique de 8 semaines sur des chiens adultes souffrant de diarrhée chronique du gros intestin. Tous les chiens ont reçu un régime thérapeutique riche en fibres, composé de diverses fibres végétales solubles et insolubles. Des échantillons de selles et de sérum ont été prélevés au début de l'étude et à plusieurs moments au cours de l'intervention. Une analyse métabolomique non ciblée a été utilisée pour établir le profil des selles et du sérum afin d'évaluer les changements au niveau de la santé gastro-intestinale et de l'activité métabolique.
Résultats. L'intervention alimentaire à base de fibres a rapidement amélioré les signes cliniques de la diarrhée. Une augmentation significative de la consistance des selles a été observée en l'espace d'une journée et la diarrhée a disparu chez la plupart des chiens en l'espace de quelques semaines. Les analyses métabolomiques ont révélé des changements significatifs au niveau des métabolites fécaux et circulants, indiquant une transition du métabolisme microbien intestinal de la fermentation des protéines (putréfaction) vers la fermentation des glucides (saccharolyse) (Figure 17). L'intervention a réduit les métabolites associés à la dégradation bactérienne des protéines et a augmenté les métabolites d'origine végétale ainsi que les postbiotiques microbiens bénéfiques, notamment les dérivés de l'indole. D'autres changements métabolomiques ont suggéré des améliorations au niveau des voies inflammatoires, du métabolisme lipidique, de l'intégrité de la barrière intestinale et des interactions hôte-microbe.
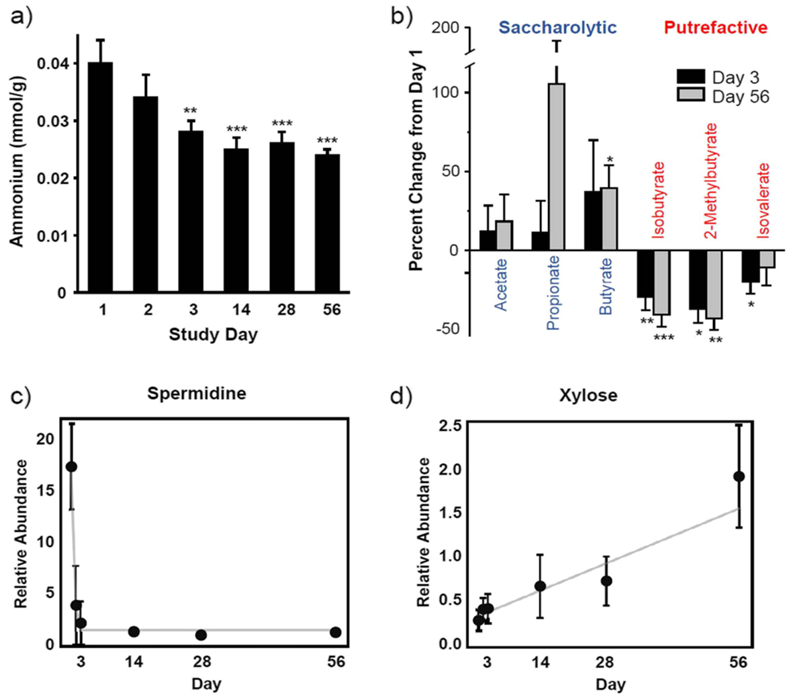
Figure 17. Impact d'une intervention alimentaire sur le métabolisme de fermentation. L'intervention à base de fibres alimentaires a réorienté le métabolisme, le faisant passer de la fermentation des protéines vers le métabolisme des glucides, comme en témoignent les baisses observées pour (A) les concentrations d'ammonium fécal (A), les acides gras à chaîne courte (B) et la spermidine (C), ainsi que l'augmentation du xylose (D). Image reproduite à partir de Fritsch et al., BMC Vet Res, 2022, sous licence CC BY 4.0.
Conclusions de l'étude
- Dans l'ensemble, les résultats de cette étude montrent que la supplémentation en fibres alimentaires modifie favorablement à la fois le métabolisme microbien et la physiologie gastro-intestinale de l'hôte, ce qui explique comment les fibres alimentaires exercent leur effet thérapeutique.
- La métabolomique a permis aux chercheurs de mesurer directement les produits métaboliques générés dans l'intestin, ce qui a fourni des informations fonctionnelles sur ces mécanismes qu'il n'aurait pas été possible d'obtenir autrement.
- Les données métabolomiques ont fourni des preuves mécanistiques de l'efficacité de l'intervention nutritionnelle, qui pourraient servir à étayer les allégations commerciales.
Optimiser la nutrition au début de la vie pour réduire l'abattage des vaches laitières
Introduction. L'alimentation en début de vie joue un rôle crucial dans le développement de la santé métabolique à long terme. Chez les veaux laitiers, une alimentation insuffisante avant le sevrage est associée à un retard de croissance, à une altération de la fonction immunitaire et à une baisse de la productivité à long terme, tandis qu'une alimentation adéquate tend à augmenter la production laitière et à favoriser une maturité reproductive précoce. Les mécanismes biologiques reliant l'alimentation en début de vie à la productivité chez les vaches adultes sont bien documentés mais mal compris.
Données préliminaires et objectifs de l'étude. Des études antérieures ont montré qu'un apport nutritionnel adéquat avant le sevrage modifie les voies métaboliques liées à l'énergie, au métabolisme des acides aminés et à la sensibilité à l'insuline, et que ces modifications persistent à l'âge adulte. Dans cette étude, les chercheurs ont émis l'hypothèse qu'un apport nutritionnel plus important avant le sevrage améliore le métabolisme énergétique et la productivité à long terme chez les vaches laitières [10]. L'objectif de cette étude était de tester cette hypothèse en déterminant comment la consommation de lait au début de la vie influence les profils métabolomiques, la production laitière et la survie au cours de plusieurs lactations.
Méthodes. 78 vaches Holstein ont été suivies de leur premier à leur cinquième vêlage. Les veaux ont été répartis de manière aléatoire dans un groupe à apport nutritionnel élevé, recevant 8 litres de lait de substitution par jour, ou dans un groupe à apport restreint, recevant 4 litres par jour jusqu’au sevrage. La croissance, les performances reproductives, la production laitière, la consommation alimentaire et la survie au cours des lactations ont été suivies. Des échantillons de sérum ont été prélevés au 60e jour de lactation et analysés par métabolomique non ciblée afin d'évaluer l'état métabolique au pic de production laitière.
Résultats. Les veaux ayant reçu des apports élevés en lait de substitution ont affiché une croissance initiale plus rapide et une meilleure sensibilité à l'insuline à un stade plus avancé de leur développement par rapport aux veaux soumis à un régime restrictif. Les vaches ayant reçu des apports élevés en lait de substitution ont également produit des rendements en matière grasse et une teneur en matière grasse plus élevés, et ont produit davantage de lait lorsque les volumes ont été corrigés en fonction des pourcentages de graisse corporelle et de protéines. Les performances de reproduction étaient similaires entre les groupes lors de la première lactation, mais lors de la deuxième lactation, les vaches ayant reçu des doses élevées de lait de substitution présentaient un taux de conception plus élevé après la première saillie que les vaches soumises à un régime restrictif. L'analyse de survie a montré que les vaches ayant reçu une alimentation plus riche au début de leur vie présentaient un risque nettement plus faible d'être réformées avant les vêlages ultérieurs (Figure 18). Le profil métabolomique au pic de lactation a révélé des différences métaboliques persistantes entre les groupes, notamment des modifications des métabolites d'acides aminés, de lipides, de nucléotides et de glucides (figure 19). Les analyses des voies métaboliques ont mis en évidence des altérations du métabolisme des purines, des pyrimidines, de la sphingosine, des acides gras et du cycle de Krebs, suggérant que la nutrition précoce influence la programmation métabolique à long terme.
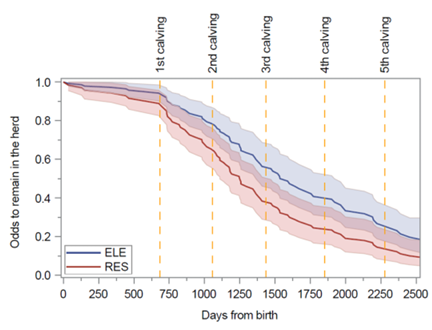
Figure 18. Probabilité que les vaches restent dans le troupeau, et échappent ainsi à l'abattage, en fonction du nombre de jours écoulés depuis la naissance pour les vaches ayant reçu un apport nutritionnel enrichi (ligne bleue) ou restreint (ligne rouge) avant le sevrage. Image reproduite à partir de Leal et al., J Dairy Sci, 2025, sous licence CC BY 4.0.
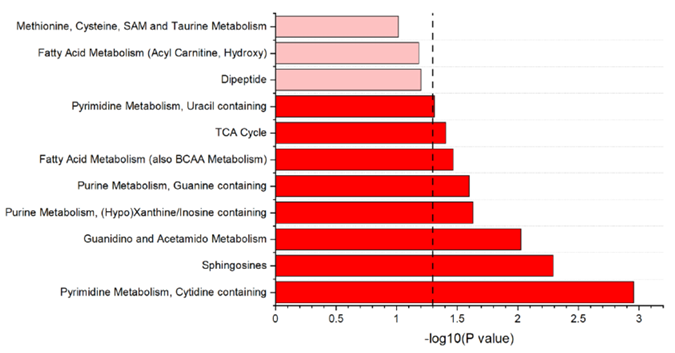
Figure 19. Représentation des voies métaboliques significatives mises en évidence par une analyse d'enrichissement basée sur les métabolites détectés dans le sérum de vaches laitières en pleine lactation. Les voies métaboliques représentées en rouge foncé ont une valeur de p < 0,05 ; celles représentées en rouge clair ont une valeur de p < 0,10. Image reproduite à partir de Leal et al., J Dairy Sci, 2025, sous licence CC BY 4.0.
Conclusions de l'étude
- Les résultats de cette étude démontrent l'existence d'un lien direct entre la nutrition au cours des premières années de vie et le fonctionnement métabolique à long terme, ce qui peut orienter les choix en matière d'alimentation animale afin d'améliorer, à terme, la productivité et la durabilité tout au long de la vie des vaches laitières, ce qui se traduira par des économies de coûts et des avantages nutritionnels pour l'homme.
Améliorer la qualité de la viande bovine en caractérisant la relation entre la qualité des sols, l'alimentation animale et la santé du bétail
Introduction. Dans l'industrie bovine américaine, les bovins sont généralement nourris avec des régimes à base de maïs pendant les derniers mois de leur vie afin d'optimiser leur croissance et la qualité de leur carcasse. La production de cultures fourragères repose sur des monocultures, des engrais synthétiques et l'irrigation, ce qui appauvrit les sols en nutriments, dégrade leur structure et contamine les sources d'eau. Il a été démontré que l'engraissement des animaux au pâturage, grâce à des pratiques de pâturage adaptatif et tournant, améliore la diversité végétale, le carbone du sol et le fonctionnement général de l'écosystème. Les animaux qui broutent du fourrage frais consomment des quantités plus importantes et plus variées de polyphénols, et l'enrichissement de ces composés dans l'alimentation du bétail peut améliorer la santé métabolique des animaux et augmenter la densité nutritionnelle globale de la viande et du lait. Afin de mieux éclairer les pratiques de pâturage, des recherches plus approfondies sont nécessaires pour établir des liens potentiels entre les indicateurs de santé des sols, des animaux et des plantes.
Données préliminaires et objectifs de l'étude. Les objectifs de cette étude étaient les suivants : 1) déterminer les différences potentielles entre les profils métaboliques de la viande bovine issue d'animaux nourris à l'herbe et ceux issus d'animaux nourris au grain, 2) évaluer les liens entre ces profils et la santé des sols ainsi que l'abondance des nutriments, et 3) analyser les biomarqueurs de santé chez les animaux afin de mieux comprendre les impacts potentiels de différents statuts nutritionnels sur leur santé et la composition nutritionnelle de leur viande [11].
Résultats. Des différences notables ont été observées en matière de santé des sols, de composition des aliments pour animaux et de profils métaboliques de la viande bovine entre les systèmes de production basés sur le pâturage et ceux nourris aux céréales. Les sols des pâturages présentaient généralement une teneur en matière organique, une disponibilité des nutriments et des teneurs en minéraux plus élevées que les sols des champs de maïs utilisés pour la production fourragère. La métabolomique non ciblée a révélé des variations biochimiques substantielles entre le fourrage de pâturage et les rations mixtes à base de céréales (figure 20). Le fourrage de pâturage contenait des teneurs plus élevées en composés phytochimiques, en composés phénoliques et en composés liés aux oméga-3, tandis que les rations à base de céréales présentaient des teneurs plus élevées en dipeptides et en métabolites liés aux oméga-6. Ces différences alimentaires se reflétaient dans la viande elle-même. Le bœuf nourri à l'herbe contenait des niveaux plus élevés de composés phénoliques, d'acides gras oméga-3, d'acylcarnitines à longue chaîne, d'antioxydants et de dérivés de la vitamine E et de la vitamine A. En revanche, le bœuf nourri aux céréales contenait des niveaux plus élevés de certains dipeptides, de sucres, de vitamines B et de marqueurs de stress oxydatif (figure 21).
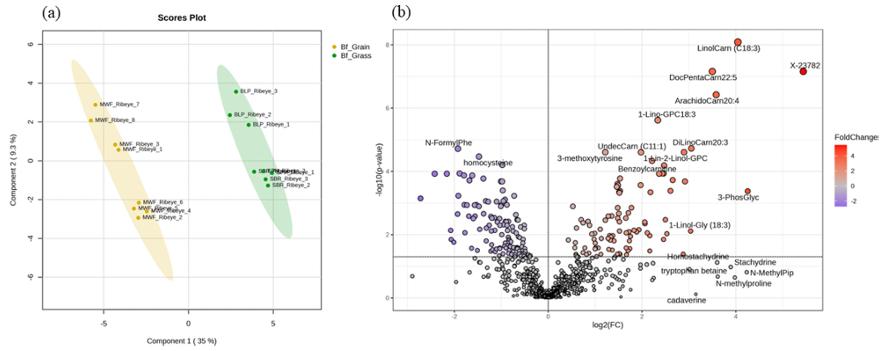
Figure 20. Analyse multivariée et univariée des profils métaboliques d'échantillons de viande bovine issue d'animaux nourris à l'herbe et d'animaux nourris au grain. (A) Analyse discriminante par moindres carrés partiels (PLS-DA) et (B) graphique en volcan des métabolites différentiels dans des échantillons de viande bovine issue d'animaux nourris à l'herbe (n = 8) et d'animaux nourris au grain (n = 8). Image reproduite à partir de Ahsin et al., NPJ Sci Food, 2025, sous licence CC BY 4.0.
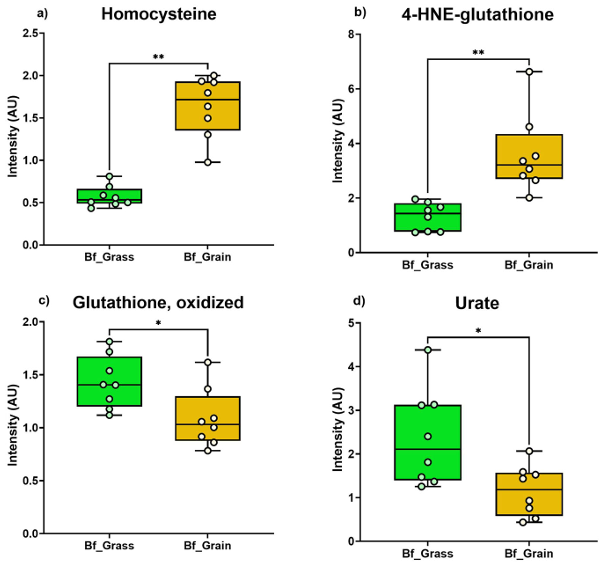
Figure 21. Marqueurs du stress oxydatif et marqueurs antioxydants dans la viande bovine issue de bovins nourris à l'herbe (Bf_Grass) et de bovins nourris au grain (Bf_Grain). Le graphique à barres présente les concentrations des marqueurs du stress oxydatif : (A) homocystine, (B) 4-HNE-glutathion, et des antioxydants : (C) glutathion oxydé, (D) urate, pour les deux groupes de bovins. Image reproduite à partir de Ahsin et al., NPJ Sci Food, 2025, sous licence CC BY 4.0.
Points à retenir de ce chapitre
- La métabolomique est un outil extrêmement précieux qui permet d'acquérir des connaissances biologiques allant au-delà de ce que peuvent offrir les autres sciences « omiques ».
- Les informations issues de la métabolomique peuvent s'avérer essentielles pour réduire les coûts et prendre l'avantage sur la concurrence.

Télécharger le guide complet au format PDF
Téléchargez ce guide complet conçu pour vous apprendre les tenants et les aboutissants de l'un des outils omiques les plus puissants de la boîte à outils de tout scientifique : la métabolomique.
Télécharger maintenantNous contacter
Parler avec un expert
Demandez un devis pour nos services, obtenez plus d'informations sur les types d'échantillons et les procédures de manipulation, demandez une lettre de soutien ou posez une question sur la façon dont la métabolomique peut faire avancer votre recherche.
Siège social
617 Davis Drive, Suite 100
Morrisville, NC 27560
Adresse postale :
P.O. Box 110407
Research Triangle Park, NC 27709
+1 (919) 572-1711
Siège international
Metabolon GmbH
Zeppelinstraße 3
85399 Hallbergmoos
Allemagne