Chapitre 4
La métabolomique au service de la recherche translationnelle
Dans ce chapitre, vous découvrirez comment la métabolomique peut contribuer à l'élaboration de modèles de maladies reflétant au mieux la biologie humaine, comment elle peut être utilisée pour caractériser de nouveaux mécanismes de progression de la maladie dans des modèles précliniques qui se transposent remarquablement bien chez les patients humains, et comment elle peut aider à expliquer pourquoi les traitements de référence d'une maladie ne sont que partiellement efficaces, ainsi que les processus biologiques à cibler pour combler cette lacune dans la prise en charge.
Vue d'ensemble
En tant que discipline « omique » la plus proche du phénotype, la métabolomique a permis de mieux comprendre les liens entre les facteurs causaux et la maladie. Contrairement aux gènes et aux protéines, les voies biochimiques sont conservées d’une espèce à l’autre. La métabolomique a donc joué un rôle déterminant dans la caractérisation des biomarqueurs et des voies pathologiques au niveau cellulaire ou dans des modèles animaux, dont les résultats sont transposables à l’être humain. Dans ce chapitre, nous aborderons des études clés dans lesquelles la métabolomique a joué un rôle essentiel pour identifier une signature biomarqueur importante ou une cible thérapeutique, caractériser un mécanisme pathologique particulièrement pertinent chez les patients humains, ou développer un nouveau modèle de maladie nettement plus proche de la physiopathologie humaine que les modèles existants. Les études de cas présentées dans ce chapitre traitent ces sujets comme suit :
- Modèles de maladies : études de cas n° 1, 2 et 4
- Biomarqueurs : études de cas n° 2, 3 et 4
- Mécanismes : Études de cas 1, 2, 3 et 4
Modèles pathologiques, biomarqueurs et mécanismes
Étude de cas n° 1 : Développement de modèles animaux de la dyslipidémie reflétant fidèlement la biologie humaine
Introduction. Les maladies cardiovasculaires (MCV) constituent l'une des principales causes de décès dans le monde, et la dyslipidémie en est un facteur de risque majeur. Bien que les statines puissent se révéler très efficaces pour réduire le taux de cholestérol LDL et diminuer le risque de MCV, de nombreux patients présentent des effets indésirables, ce qui souligne la nécessité de trouver de nouvelles cibles thérapeutiques. Cependant, les efforts visant à mettre au point de nouveaux traitements n'ont donné lieu qu'à un succès clinique limité, souvent attribué à des modèles animaux précliniques inadéquats qui ne reflètent pas fidèlement le métabolisme lipidique humain.
Données préliminaires et objectifs de l'étude. Afin de pallier les limites des modèles animaux actuels du métabolisme lipidique, les chercheurs ont eu recours à la métabolomique pour analyser et comparer les profils lipidiques plasmatiques chez plusieurs espèces animales et chez l'homme, dans le but de déterminer quels modèles se rapprochent le plus de la dyslipidémie humaine, tant à l'état basal qu'en réponse à un traitement par statines [12].
Méthodes. Plusieurs souches de souris, d’autres espèces non primates et des primates non humains ont été analysés parallèlement à des sujets humains présentant une dyslipidémie. Les animaux ont été nourris soit avec une alimentation standard de laboratoire, soit avec un régime riche en graisses ou en cholestérol. La plupart des espèces ont reçu un traitement à la simvastatine ou un placebo pendant deux semaines afin de permettre une comparaison entre les profils lipidiques de base et ceux observés en réponse au médicament. Les échantillons de plasma ont été analysés pour déterminer les lipides et les lipoprotéines à l'aide de la métabolomique. Les lipides ont également été mesurés à l'aide de mesures cliniques traditionnelles, notamment le cholestérol total, le LDL, le HDL et les triglycérides. Des méthodes basées sur la distance ont été utilisées pour quantifier le degré de similitude entre chaque modèle animal et la dyslipidémie humaine pour plusieurs fractions lipidiques.
Résultats. Des différences notables ont été observées dans les profils lipidiques plasmatiques de base entre les différents modèles animaux, seul un sous-groupe présentant une forte similitude avec les humains atteints de dyslipidémie. Les primates non humains ont montré la plus grande similitude avec les humains en ce qui concerne le cholestérol total, l'équilibre LDL/HDL et la distribution globale des lipoprotéines (figure 22). De nombreux modèles couramment utilisés, notamment les souris, les lapins et les rats, présentaient des différences significatives par rapport aux humains. Par exemple, de nombreux modèles de rongeurs transportaient la majeure partie du cholestérol dans les HDL (un profil athéroprotecteur), contrairement aux humains, tandis que les primates non humains et certains modèles modifiés, tels que les souris exprimant la CETP, présentaient des profils lipidiques plus proches de ceux des humains. La composition en acides gras des principales classes de lipides était globalement conservée chez certaines espèces, mais divergeait chez d’autres. Chez l'humain, la plupart des acides gras provenaient des voies non essentielles et oméga-6, et ce profil était le mieux reproduit chez les primates non humains, les chiens et quelques modèles de rongeurs. En comparant la composition lipidique globale des huit principales fractions lipidiques, les modèles de primates non humains se sont révélés les plus proches de l'humain (Figure 23). En réponse au traitement par simvastatine, les primates non humains et les chiens ont présenté des modifications lipidiques les plus cohérentes avec celles de l'humain, tandis que la plupart des modèles de rongeurs ont montré des réponses faibles, voire opposées.
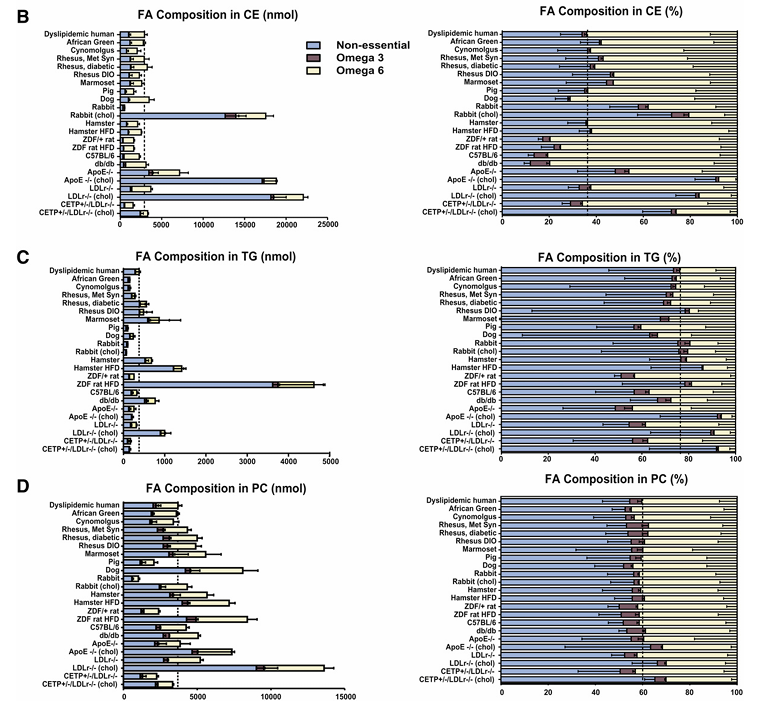
Figure 22. Composition en acides gras (AG) des esters de cholestérol (EC), des triglycérides (TG) et de la phosphatidylcholine (PC) plasmatiques chez différentes espèces animales. Les AG sont classés en trois catégories : les AG non essentiels (bleu clair), les AG de la voie oméga-3 (marron) et les AG de la voie oméga-6 (jaune clair). Les AG mesurés ont été additionnés pour chaque voie. Le panneau de gauche indique les quantités absolues (nmol) d'AG dans chaque voie et le panneau de droite indique les quantités relatives (%) pour les CE plasmatiques, (A) les TG (B) et (C) les PC. Image reproduite à partir de Yin et al., J Lipid Res, 2012, sous licence CC BY 4.0.
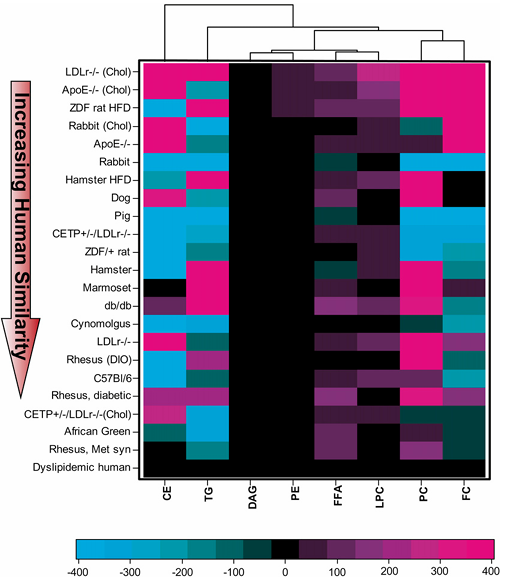
Figure 23. Comparaison sous forme de dendrogramme de la similitude des lipides plasmatiques de base, sur la base de huit fractions lipidiques circulantes principales (CE, TG, DAG, PE, FFA, LPC, PC et FC) entre les espèces. La différence entre les moyennes de chaque fraction lipidique donnée pour chaque modèle animal et pour un humain dyslipidémique a été calculée et pondérée en fonction de la proportion de cette fraction lipidique dans le total des lipides chez l'humain. La distance pondérée globale de chaque modèle animal par rapport aux humains dyslipidémiques a été calculée, et les modèles ont été classés par distance. Image reproduite à partir de Yin et al., J Lipid Res, sous licence CC BY 4.0.
Conclusions de l'étude
- Cette étude montre que les profils lipidiques de base et ceux observés après traitement sont essentiels pour évaluer dans quelle mesure les modèles animaux reflètent la maladie chez l'homme.
- Les primates non humains constituent les modèles les plus représentatifs de la dyslipidémie humaine, et les chiens présentent des caractéristiques similaires en ce qui concerne la réponse aux médicaments. En revanche, de nombreux modèles animaux couramment utilisés, notamment les souris ApoE-/- et LDLr-/- , présentaient des différences significatives par rapport à l'être humain.
- Non seulement la métabolomique a permis une comparaison exhaustive des profils lipidiques entre plusieurs modèles animaux et l'être humain, mais elle a également démontré que l'association d'un profil lipidique complet (lipidomique) avec des données fonctionnelles sur la réponse aux médicaments offre un cadre plus précis pour évaluer la similitude des maladies et améliorer la validation des cibles.
Étude de cas n° 2 : Caractérisation des mécanismes de la maladie de Niemann-Pick qui reflètent le phénotype des patients humains
Introduction. La maladie de Niemann-Pick de type C1 (NPC1) est une affection neurodégénérative rare et évolutive, principalement causée par des mutations du gène NPC1 qui entraînent une altération du transport intracellulaire du cholestérol et une accumulation de lipides dans les lysosomes. La maladie se manifeste généralement pendant l'enfance par une ataxie et un déclin cognitif ; en l'absence d'intervention rapide, les patients en meurent souvent à l'adolescence. L'un des principaux défis dans la prise en charge de la maladie de NPC1 est l'absence de test diagnostique simple et non invasif, ce qui peut retarder le diagnostic de plusieurs années.
Données préliminaires et objectifs de l'étude. La maladie NPC1 est étroitement associée au stress oxydatif, comme le démontrent l'augmentation des espèces réactives de l'oxygène (ERO) et de la peroxydation lipidique observées dans des modèles cellulaires et animaux, ainsi que la diminution de la capacité antioxydante chez les patients. L'environnement oxydatif, associé à un excès de cholestérol cellulaire, favorise la formation de produits d'oxydation du cholestérol (oxystérols). Sur la base de ces résultats précédemment rapportés, les chercheurs de cette étude ont émis l'hypothèse que les produits d'oxydation du cholestérol circulants pourraient servir de biomarqueurs sanguins sensibles et spécifiques de la maladie NPC1 [13]. L'objectif de cette étude était de déterminer si les oxystérols plasmatiques permettaient de distinguer les patients atteints de la maladie NPC1 des individus sains et si les taux d'oxystérols étaient corrélés à la gravité de la maladie.
Méthodes. Des souris Npc1-/- et un modèle félin de la maladie NPC1 ont été utilisés pour évaluer les oxystérols en tant que biomarqueurs de la maladie NPC1. Des échantillons de plasma, de tissu hépatique et cérébral, ainsi que de liquide céphalo-rachidien (LCR) ont été prélevés sur les animaux tout au long de leur vie. Parallèlement, des échantillons de plasma, de LCR et de fibroblastes humains ont été prélevés chez des patients atteints de la maladie NPC1, des témoins sains, des porteurs hétérozygotes d'une mutation du gène NPC1 et des personnes atteintes d'autres maladies de surcharge lysosomale afin d'évaluer la spécificité des biomarqueurs. Les oxystérols ont été mesurés à l'aide d'une métabolomique ciblée. Les différences dans les taux d'oxystérols et leur corrélation avec la gravité de la maladie ont été déterminées à l'aide d'une analyse de variance (ANOVA) et d'analyses de corrélation.
Résultats. Plusieurs oxystérols présentaient des taux significativement élevés dans le plasma et les tissus des souris Npc1-/- ; une augmentation modérée a été observée avant l'apparition des symptômes neurologiques, puis s'est accentuée à mesure que la maladie progressait. Chez l'homme, les oxystérols non enzymatiques 3β,5α,6β-triol et 7-cétocolestérol (7-KC) étaient significativement plus élevés chez les patients atteints de NPC1 que chez les témoins et les porteurs hétérozygotes, ce qui a permis de détecter les cas avec une sensibilité et une spécificité élevées. Ces biomarqueurs ont clairement permis de distinguer les patients atteints de NPC1 des individus sains et de ceux atteints d'autres maladies de surcharge lysosomale (Figure 24). Des concentrations plus élevées de 3β,5α,6β-triol et de 7-KC étaient associées à une apparition plus précoce de la maladie et à une plus grande gravité de celle-ci, et un indice combinant ces marqueurs a amélioré la capacité prédictive. Dans le modèle félin de la NPC1, le traitement par cyclodextrine a réduit de manière significative les taux d'oxystérols, démontrant que ces biomarqueurs peuvent également servir à suivre la réponse thérapeutique (Figure 25).
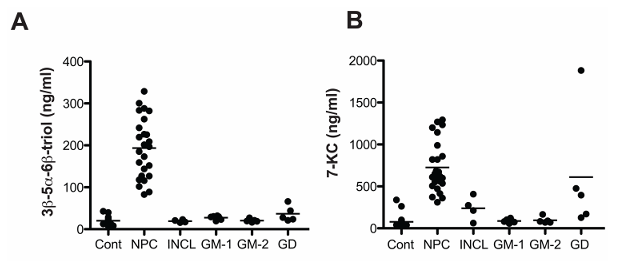
Figure 24. Comparaison des concentrations plasmatiques d'oxystérols dans la maladie NPC1 et d'autres maladies de surcharge lysosomale. Concentrations de (A) 3β,5α,6β-triol et (B) 7-KC dans des échantillons de plasma à jeun provenant de sujets témoins, atteints de la maladie NPC1, de la lipofuscinoses céroïde neuronale infantile (INCL), de la gangliosidose GM-1 (GM-1), de la gangliosidose GM-2 (GM-2) et de la maladie de Gaucher (GD). Image reproduite à partir de Proter et al., Sci Transl Med, 2010, sous licence CC BY 4.0.
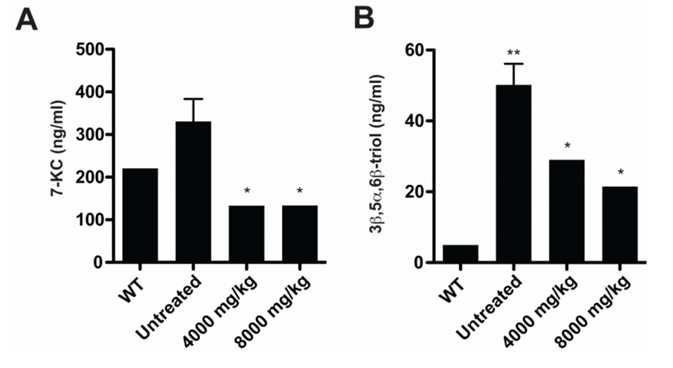
Figure 25. Les biomarqueurs des oxystérols circulants diminuent en réponse au traitement par cyclodextrine. (A) Les concentrations sériques de 7-KC et (B) de 3β,5α,6β-triol ont été mesurées chez des chats WT (4 à 16 semaines) et NPC1 (16 semaines) non traités, ainsi que chez des chats NPC1 (16 à 18 semaines) ayant reçu une injection sous-cutanée unique de 4 000 ou 8 000 mg/kg à 3 semaines (n = 2 à 4 par groupe). Image reproduite à partir de Proter et al., Sci Transl Med, 2010, sous licence CC BY 4.0
Conclusions de l'étude
- Cette étude démontre que les oxystérols constituent des biomarqueurs hautement sensibles et spécifiques de la maladie NPC1, ce qui répond à un besoin important non satisfait en matière de diagnostic et de suivi de cette pathologie.
- Contrairement à d'autres maladies où l'augmentation des taux d'oxystérols est modérée, la NPC1 se caractérise par des élévations importantes et spécifiques à la maladie, qui permettent de la distinguer clairement des témoins et d'autres pathologies.
- Étant donné que les oxystérols sont corrélés à la gravité de la maladie et à l'âge d'apparition de celle-ci, ils pourraient également s'avérer utiles pour suivre l'évolution de la maladie. La baisse observée des taux d'oxystérols à la suite d'un traitement dans des modèles animaux suggère qu'ils pourraient servir de critères de substitution dans les essais cliniques afin d'aider à évaluer l'efficacité thérapeutique.
Étude de cas n° 3 : Caractérisation des interactions entre l'intestin et le cerveau propres aux personnes atteintes de troubles du spectre autistique
Introduction. À l'heure actuelle, les troubles du spectre autistique (TSA) sont diagnostiqués sur la base de critères comportementaux plutôt que de marqueurs biologiques, ce qui contribue à retarder le diagnostic et limite les possibilités d'intervention précoce. La prévalence croissante des TSA et l'absence de traitements approuvés par la FDA pour les symptômes principaux rendent nécessaire la mise au point de biomarqueurs moléculaires objectifs afin de permettre une détection plus précoce et de soutenir des thérapies personnalisées. Les TSA sont de plus en plus associés à des anomalies métaboliques, notamment un dysfonctionnement mitochondrial et un stress oxydatif. Cependant, les résultats des études antérieures ont été incohérents en raison de la petite taille des échantillons, de la couverture limitée des métabolites et de la variabilité dans la conception des études, ce qui a entravé les tentatives d'identification de signatures métaboliques cohérentes. Une complexité supplémentaire provient de l'alimentation et du microbiome intestinal, qui peuvent influencer de manière significative les profils métaboliques et interagir avec des facteurs de risque génétiques pour influencer la biologie des TSA.
Données préliminaires et objectifs de l'étude. Afin de combler ces lacunes dans les connaissances, les chercheurs ont cherché à réaliser des analyses métabolomiques approfondies d'échantillons de plasma et de selles provenant d'une vaste cohorte d'enfants atteints de TSA, ainsi que d'un groupe témoin neurotypique [14]. L'objectif de cette étude était d'identifier des signatures métaboliques fiables associées aux TSA et de caractériser les liens entre le métabolisme et les symptômes comportementaux.
Méthodes. Les participants étaient âgés de 3 à 12 ans. Les diagnostics de TSA ont été confirmés à l'aide d'outils cliniques standardisés, et les témoins neurotypiques ont fait l'objet d'une sélection afin de garantir un développement typique. Une partie des participants a également subi une évaluation des symptômes gastro-intestinaux (GI) afin de répartir les personnes atteintes de TSA en deux groupes (avec et sans dysfonctionnement GI) en vue d'analyses complémentaires. Le plasma et les selles ont été analysés à l'aide d'une métabolomique globale et d'un profil lipidique complet. Les différences métaboliques entre les groupes d'étude ont été déterminées à l'aide de comparaisons basées sur l'ANOVA. L'apprentissage automatique de type « Random Forest » a été utilisé pour déterminer si les profils métaboliques permettaient de distinguer les patients atteints de TSA des témoins. De plus, le microbiote fécal de donneurs atteints de TSA et de donneurs neurotypiques a été transplanté chez des souris germ-free afin d'examiner si les signatures métaboliques associées au microbiome pouvaient être transférées et de mieux comprendre les interactions intestin-cerveau.
Résultats. Des centaines de métabolites présentaient des différences entre les groupes, et les profils métaboliques permettaient de distinguer modestement les personnes atteintes de TSA des individus neurotypiques, les principaux métabolites discriminants comprenant des lipides, des hormones stéroïdes et des composés d'origine microbienne. Des modifications des taux d'acyl-carnitines et de métabolites liés à l'énergie suggéraient une altération du métabolisme énergétique cellulaire ; on a également observé des perturbations dans les voies métaboliques des acides aminés et du glutathion, compatibles avec un stress oxydatif accru (figure 26). Bon nombre de ces voies métaboliques altérées étaient corrélées aux scores de gravité comportementale, établissant ainsi un lien entre les anomalies métaboliques et les caractéristiques cliniques de l'autisme. Plusieurs métabolites d'origine microbienne, dont le sulfate de 4-éthylphényle, présentaient des taux élevés chez les personnes atteintes d'autisme, et certains pouvaient être transférés à des souris par transplantation de microbiote fécal, ce qui suggère une contribution causale des microbes intestinaux aux changements métaboliques (Figure 27).
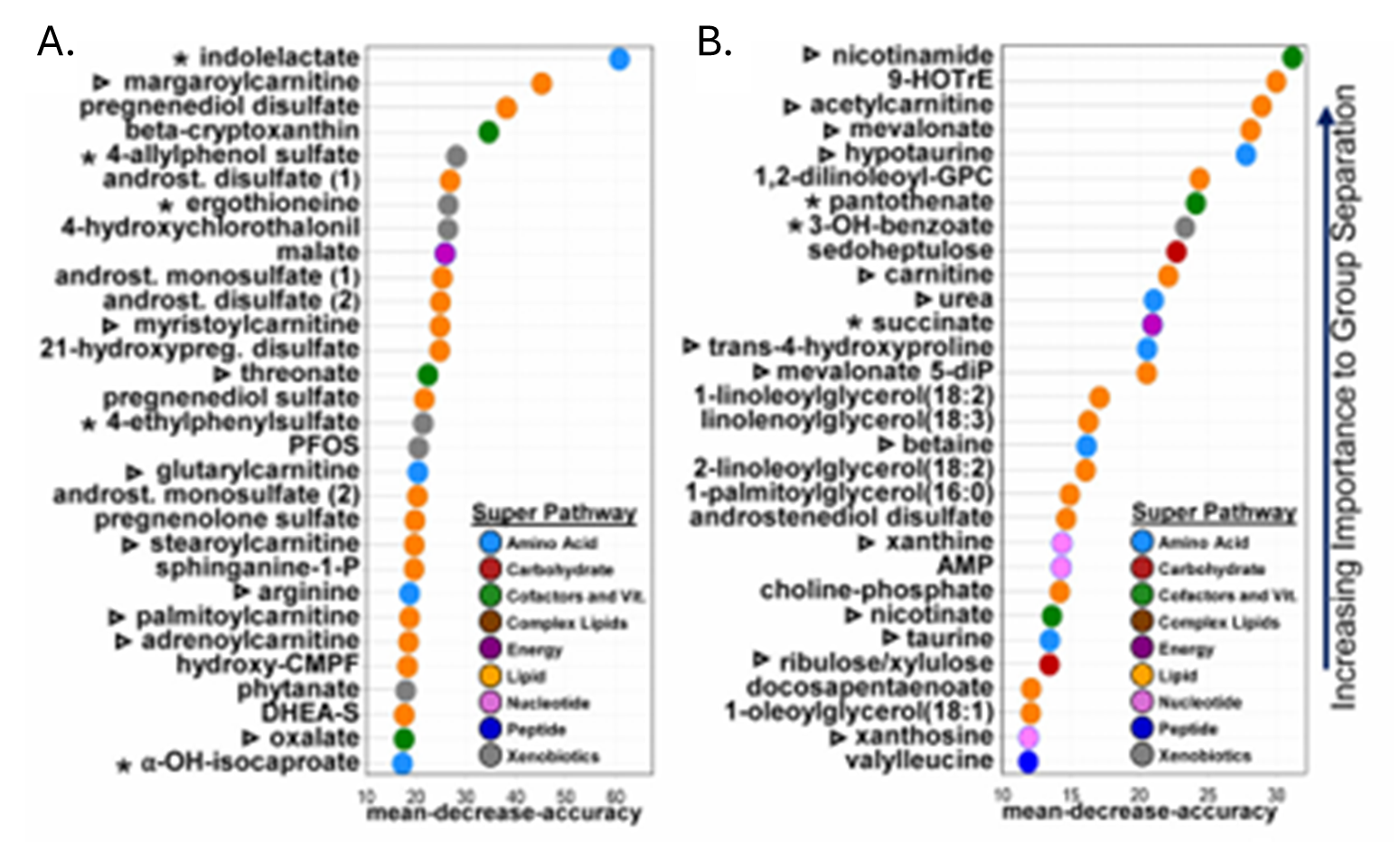
Figure 26. Les métabolomes plasmatiques et fécaux diffèrent entre les groupes atteints de TSA et les groupes neurotypiques. Les 30 métabolites les plus distinctifs entre chaque groupe dans le plasma (A) et les selles (B), selon une analyse par forêt aléatoire, avec la diminution moyenne de la précision représentée sur l'axe des x. Image reproduite à partir de Needham et al., Biol Psychiatry, 2021, sous licence CC BY 4.0
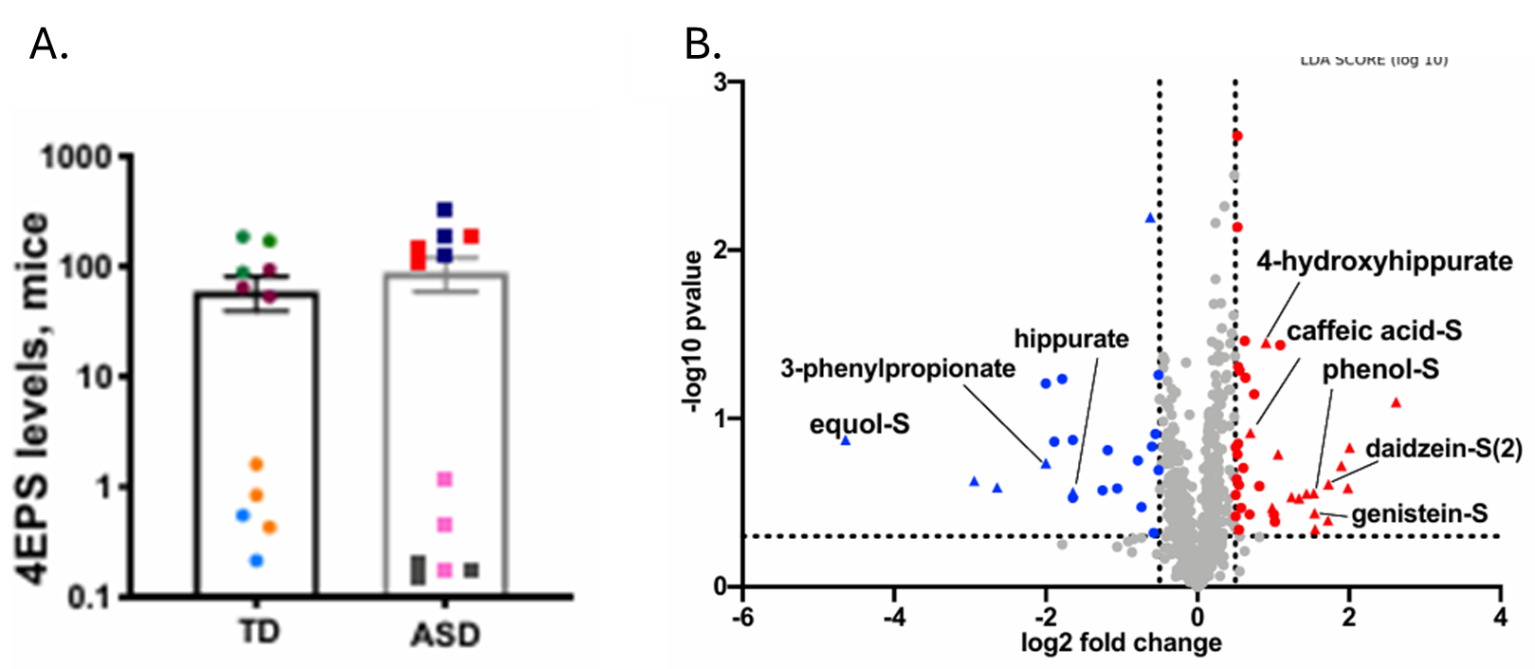
Figure 27. Transfert de microbiote fécal humain chez la souris. (A) Valeurs d’intensité normalisées indiquant les niveaux relatifs de sulfate de 4-éthylphényle (4EPS) chez des souris colonisées par des donneurs neurotypiques ou atteints de TSA, colorées en fonction du donneur. (B) Valeurs d’intensité normalisées indiquant les niveaux relatifs de 4EPS chez des souris colonisées par des donneurs neurotypiques ou atteints de TSA, colorées en fonction du donneur. Image reproduite à partir de Needham et al., Biol Psychiatry, 2021, sous licence CC BY 4.0.
Conclusions de l'étude
- Cette étude a mis en évidence des profils métaboliques variés associés à des symptômes gastro-intestinaux et à des scores comportementaux, ce qui suggère que les TSA ne peuvent s'expliquer par un seul mécanisme, mais qu'ils sont plutôt influencés par le risque génétique, l'alimentation et le microbiome intestinal.
- De nombreux métabolites modifiés provenaient des microbes intestinaux ou étaient influencés par ceux-ci. Il a été démontré que certaines de ces signatures métaboliques pouvaient être transférées à des souris par le biais d'une transplantation de microbiote, ce qui suggère que l'intestin et le système immunitaire peuvent influencer le fonctionnement du cerveau par l'intermédiaire des métabolites circulants.
- Dans l'ensemble, cette étude suggère que l'intégration de la métabolomique aux données cliniques et environnementales pourrait améliorer le diagnostic des TSA et permettre d'identifier de nouvelles cibles thérapeutiques.
Étude de cas n° 4 : Mise en lumière d'un nouveau mécanisme expliquant pourquoi les traitements de référence de la lymphangioleiomyomatose sont partiels et réversibles
Introduction. La lymphangioleiomyomatose (LAM) est une maladie pulmonaire rare et évolutive qui touche principalement les femmes et se caractérise par une prolifération anormale de cellules de type musculaire lisse, entraînant une destruction kystique des poumons et une insuffisance respiratoire. La LAM est liée à des mutations des gènes du complexe de la sclérose tubéreuse (TSC1/TSC2), qui entraînent une hyperactivation de la voie de signalisation mTORC1, un régulateur clé de la croissance cellulaire et du métabolisme. Cependant, l'activation de mTORC1 à elle seule n'explique pas entièrement la pathogenèse de la LAM, car les bénéfices cliniques des inhibiteurs de mTOR, tels que la rapamycine, sont incomplets et réversibles. La prédominance frappante de la LAM chez les femmes suggère un rôle de l'estradiol dans la progression de la maladie.
Données préliminaires et objectifs de l'étude. Des études antérieures ont montré que l'estradiol peut favoriser la croissance tumorale et les métastases dans des modèles présentant une déficience en TSC2, ce qui suggère une implication hormonale dans la biologie de la maladie. De plus, des voies impliquant des médiateurs inflammatoires ont également été identifiées comme des médiateurs importants de la biologie de la maladie. Les chercheurs ont émis l'hypothèse que la biosynthèse des prostaglandines, régulée par l'enzyme COX-2, pourrait constituer une voie en aval clé reliant la signalisation des œstrogènes et la perte de TSC2. L'objectif de cette étude était de déterminer comment l'estradiol, la déficience en TSC2 et la signalisation mTOR interagissent pour réguler la COX-2 et la production de prostaglandines, et si cette voie pourrait servir de cible thérapeutique dans la LAM [15].
Méthodes. Des études mécanistiques ont été menées à l'aide de lignées cellulaires déficientes en TSC2 et de contrôles avec « réintroduction » de TSC2. Les cellules ont été traitées avec de l'estradiol et divers inhibiteurs de voies de signalisation. Les paramètres évalués, notamment l'expression de la COX-2, l'activation des voies de signalisation et la production de prostaglandines, ont été mesurés par immunoblotting, ELISA et RT-PCR. Une inhibition génique (par exemple, shRNA Rictor) a été utilisée pour évaluer le rôle de la signalisation mTORC2. Une analyse métabolomique globale a permis d'identifier des modifications des prostaglandines et des métabolites lipidiques associés à la suite d'un traitement à l'estradiol. Des cellules déficientes en TSC2 et des cellules restaurées en TSC2 ont été implantées chez des souris Tsc+/- afin d'évaluer la croissance tumorale, la production de prostaglandines et les réponses au traitement. Les souris ont été traitées à l'aspirine ou au célécoxib afin d'évaluer l'impact thérapeutique de l'inhibition de la COX-2. Le tissu pulmonaire, le sérum, l'urine et le condensat de l'air expiré de patients atteints de LAM ont été analysés afin de valider les résultats et d'évaluer la pertinence de l'activité de la COX-2 et de la signalisation des prostaglandines dans la maladie humaine.
Résultats. L'estradiol a considérablement accru la production de prostaglandines dans les cellules déficientes en TSC2, révélant une signature métabolique distincte caractérisée par des taux accrus de prostaglandines et de métabolites associés. Dans les cellules déficientes en TSC2, les voies de signalisation MAPK et PI3K–Akt ont été activées par l'estradiol, ce qui a entraîné une augmentation de l'expression de la COX-2, suggérant ainsi un lien mécanistique entre la signalisation œstrogénique et le métabolisme lipidique inflammatoire dans la LAM. L'augmentation de l'expression de la COX-2 et de la biosynthèse des prostaglandines s'est produite indépendamment de la signalisation mTORC1. Il est intéressant de noter que la rapamycine a inhibé l'activité de mTORC1 sans pour autant réduire les taux de COX-2 ni la production de prostaglandines, tandis que l'inhibition de mTORC2 a considérablement réduit l'expression de la COX-2 et la signalisation en aval. Dans des modèles murins, l'inhibition pharmacologique de la COX-2 par l'aspirine ou le célécoxib a réduit les taux de prostaglandines, freiné la croissance tumorale et augmenté l'apoptose dans des modèles précliniques (Figure 28). Dans des échantillons humains de LAM, l'expression de la COX-2 et les prostaglandines circulantes étaient élevées, et le traitement à l'aspirine a augmenté les taux de médiateurs lipidiques anti-inflammatoires, ce qui a réduit la prolifération cellulaire.
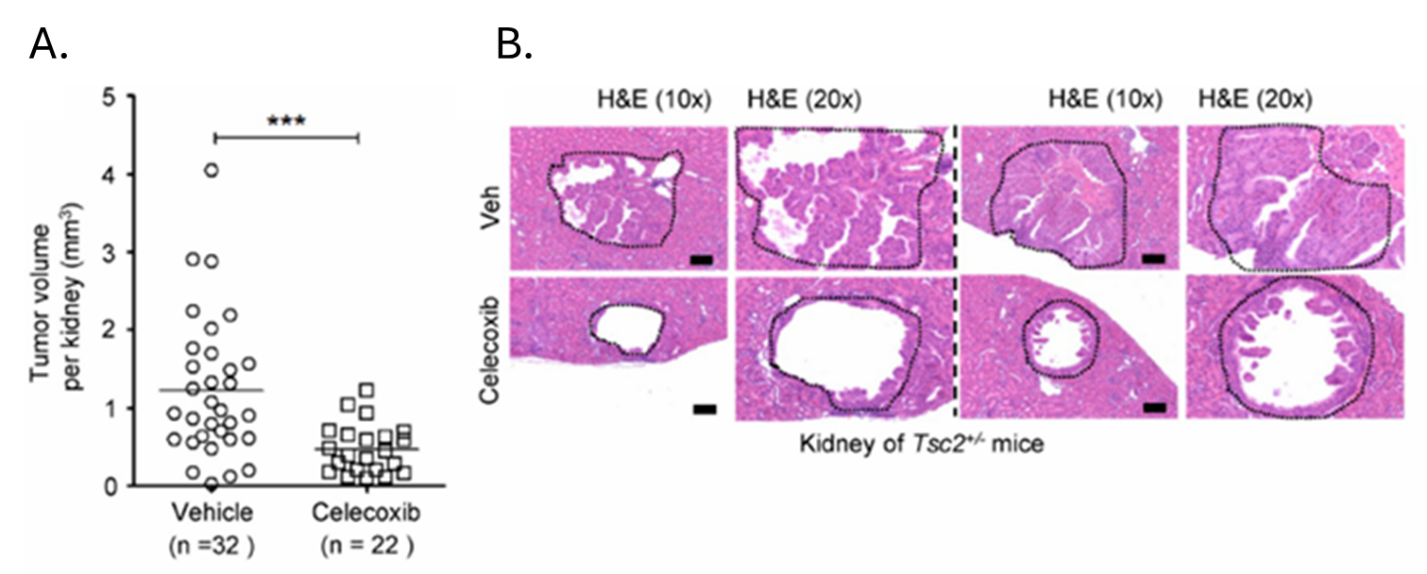
Figure 28. L'inhibition de la COX-2 empêche la survenue de tumeurs rénales et freine la progression de tumeurs xénogreffées issues de cellules déficientes en TSC2 dans des modèles précliniques. Des souris Tsc2+/2 ont été traitées soit avec un véhicule, soit avec du célécoxib (0,1 % dans la nourriture pour souris) pendant un mois, puis sacrifiées à la fin du traitement pour analyse. L'histologie des cystadénomes rénaux et les scores tumoraux rénaux microscopiques ont été évalués. (A) Les scores tumoraux rénaux microscopiques sont représentés sur une échelle linéaire (P = 0,0002). Les données proviennent de 16 groupes traités par le véhicule et de 11 groupes traités par le célécoxib. (B) Deux cystadénomes sont présentés. Les résultats sont représentatifs de 11 ou 16 souris par groupe. Image reproduite à partir de Li et al., J Exp Med, 2014, sous licence CC BY 4.0.
Conclusions de l'étude
- Cette étude a mis en évidence un nouveau lien entre la signalisation des œstrogènes, la perte de TSC2 et le métabolisme des prostaglandines dans la LAM.
- Les traitements inhibant la COX-2 ont freiné la croissance tumorale et favorisé l'apoptose dans un modèle murin de LAM. Une diminution de la prolifération cellulaire a également été observée en réponse à des traitements inhibiteurs de la COX-2 dans des cellules humaines prélevées chez des patients atteints de LAM.
- Alors qu'on pensait auparavant que la LAM était principalement induite par l'activation de mTORC1, ces résultats ont montré que la production de prostaglandines médiée par la COX-2 est un mécanisme indépendant de mTORC1 qui contribue à la progression de la maladie, ce qui explique pourquoi les traitements de référence, tels que la rapamycine, n'apportent que des bénéfices cliniques partiels.
Points à retenir de ce chapitre
- De par sa position de discipline la plus proche du phénotype, la métabolomique s'avère particulièrement utile pour caractériser les mécanismes biologiques et identifier des biomarqueurs ainsi que des cibles thérapeutiques dans des modèles précliniques qui se transposent remarquablement bien à l'être humain.
- Le recours à la métabolomique dans les études translationnelles pourrait améliorer le taux de réussite des essais cliniques, en particulier pour les pathologies pour lesquelles de nombreux traitements ont été proposés mais ont finalement échoué lors des essais cliniques en raison de l'absence de modèles translationnels fiables.

Télécharger le guide complet au format PDF
Téléchargez ce guide complet conçu pour vous apprendre les tenants et les aboutissants de l'un des outils omiques les plus puissants de la boîte à outils de tout scientifique : la métabolomique.
Télécharger maintenantNous contacter
Parler avec un expert
Demandez un devis pour nos services, obtenez plus d'informations sur les types d'échantillons et les procédures de manipulation, demandez une lettre de soutien ou posez une question sur la façon dont la métabolomique peut faire avancer votre recherche.
Siège social
617 Davis Drive, Suite 100
Morrisville, NC 27560
Adresse postale :
P.O. Box 110407
Research Triangle Park, NC 27709
+1 (919) 572-1711
Siège international
Metabolon GmbH
Zeppelinstraße 3
85399 Hallbergmoos
Allemagne